Direct ETTIN-auxin interaction controls chromatin states in gynoecium development
- Department of Crop Genetics, John Innes Centre, Norwich Research Park, United Kingdom
- Centre for Plant Sciences, Faculty of Biological Sciences, University of Leeds, United Kingdom
- Institute of Science and Technology, Austria
Figures
Figure 1 with 2 supplements
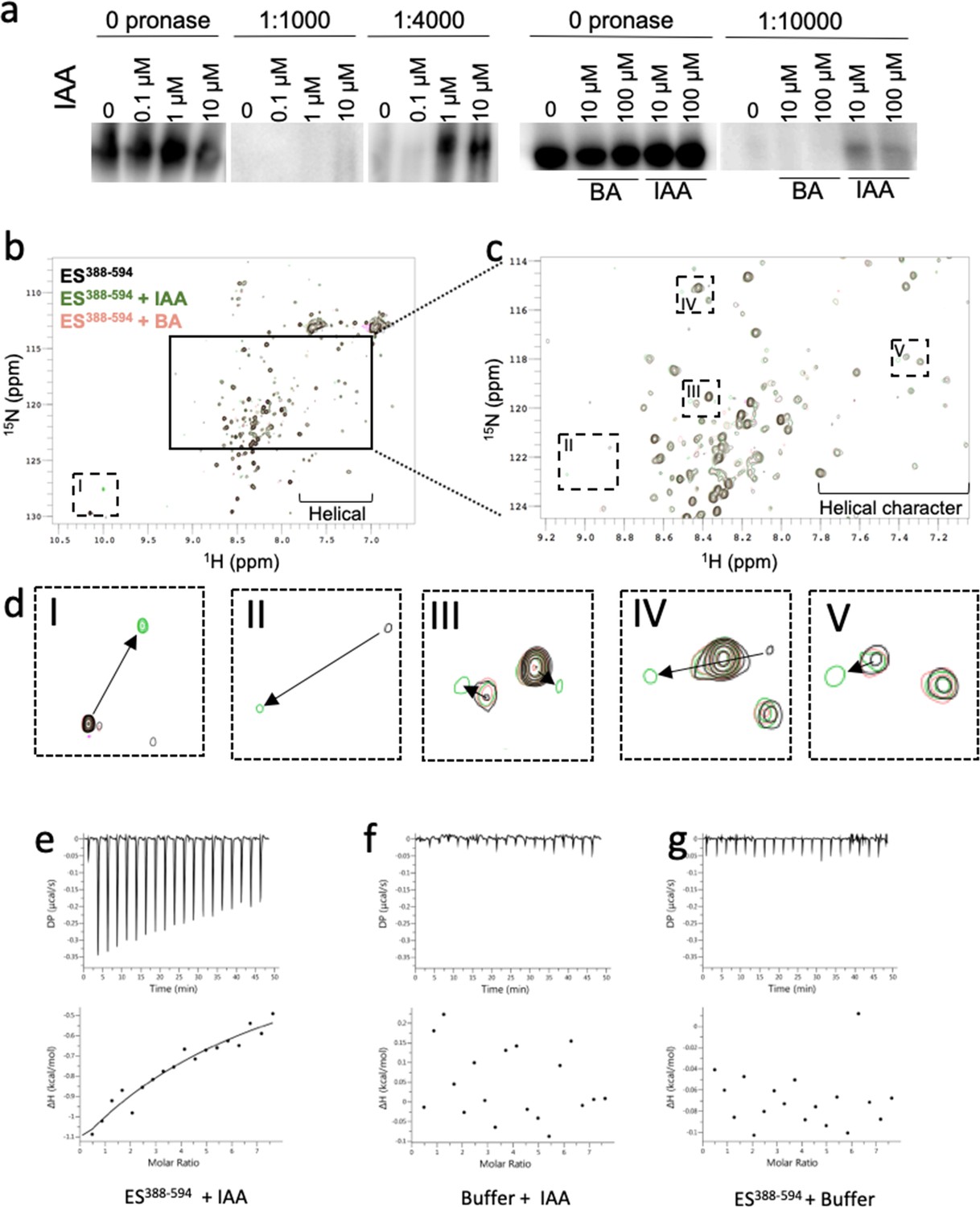
ETT directly binds auxin (IAA).
(a) DARTS assay suggests that ETT binds IAA. Seedlings from an inducible two-component ETT-FLAG line were used for the protein isolation. Samples were treated with DMSO (mock), IAA and BA and digested by different concentrations of Pronase. Samples were further analysed by western blot with an anti-FLAG antibody. (b) HSQC-NMR performed with ES388-594 protein either alone (black), with indole-3-acetic acid (IAA, green) or benzoic acid (BA, pink). ES388-594: ligand, 1:10, 50 µM:500 µM. (c) Zoom-in of the indicated rectangular region in a. (d) Zoom-in of the specific shifts (labelled I-V) in the indicated dotted rectangles in b and c. Changes in chemical shifts are indicated by arrows from control to IAA treatment. (e–g) ITC spectre showing heat exchange between ES388-594 protein and IAA (e), but not in controls (f, g). See Figure 1—figure supplement 1 for parameters used in the HSQC-NMR experiment.
-
Figure 1—source data 1
Parameters for HSQC NMR.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig1-data1-v2.xlsx
Figure 1—figure supplement 1
Original western blot images for DARTS assays.
The red boxes indicate the regions used in Figure 1a.
Figure 1—figure supplement 2

Parameters for HSQC NMR experiment.
Figure 2 with 1 supplement
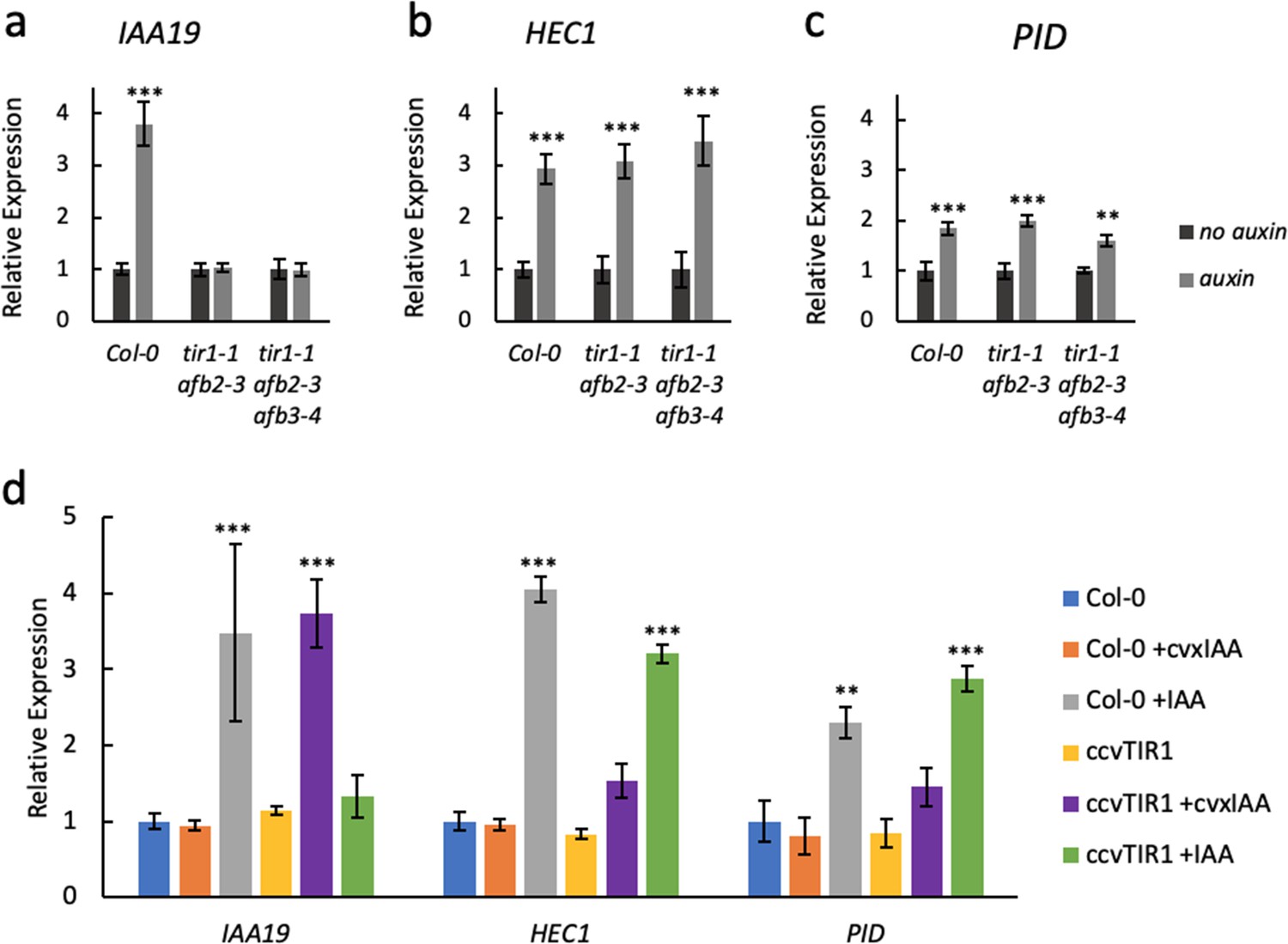
ETT regulates target gene expression independently of TIR1/AFB auxin receptors.
Expression of the canonical auxin responsive IAA19 gene (a) and the ETT-target genes HEC1 (b) and PID (c) in control-treated (dark grey) or 100 µM IAA-treated (light grey) gynoecia assayed using qRT-PCR. (a) IAA19 expression is up-regulated in response to auxin in wild-type gynoecia (Col-0) but not in tir1/afb double and triple mutants. The ETT-target genes HEC1 and PID are up-regulated in response to auxin in both wild-type and auxin receptor mutants (b c). This suggests a TIR1/AFB independent regulation of these genes. (d) Expression of IAA19, HEC1 and PID in response to treatment with 100 µM IAA and 100 µM cvxIAA in wild-type (Col-0) and pTIR1:ccvTIR1 gynoecia in the tir1 afb2 double mutant (ccvTIR1). The data confirm TIR1/AFB independent regulation of HEC1 and PID in the gynoecium. ***p<0.0001; Shown are mean ± standard deviation of three biological replicates. See Figure 2—source data 2 for statistical analyses.
-
Figure 2—source data 1
Oligonucleotides used in this study.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig2-data1-v2.xlsx
-
Figure 2—source data 2
Output of statistical tests.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig2-data2-v2.xlsx
Figure 2—figure supplement 1
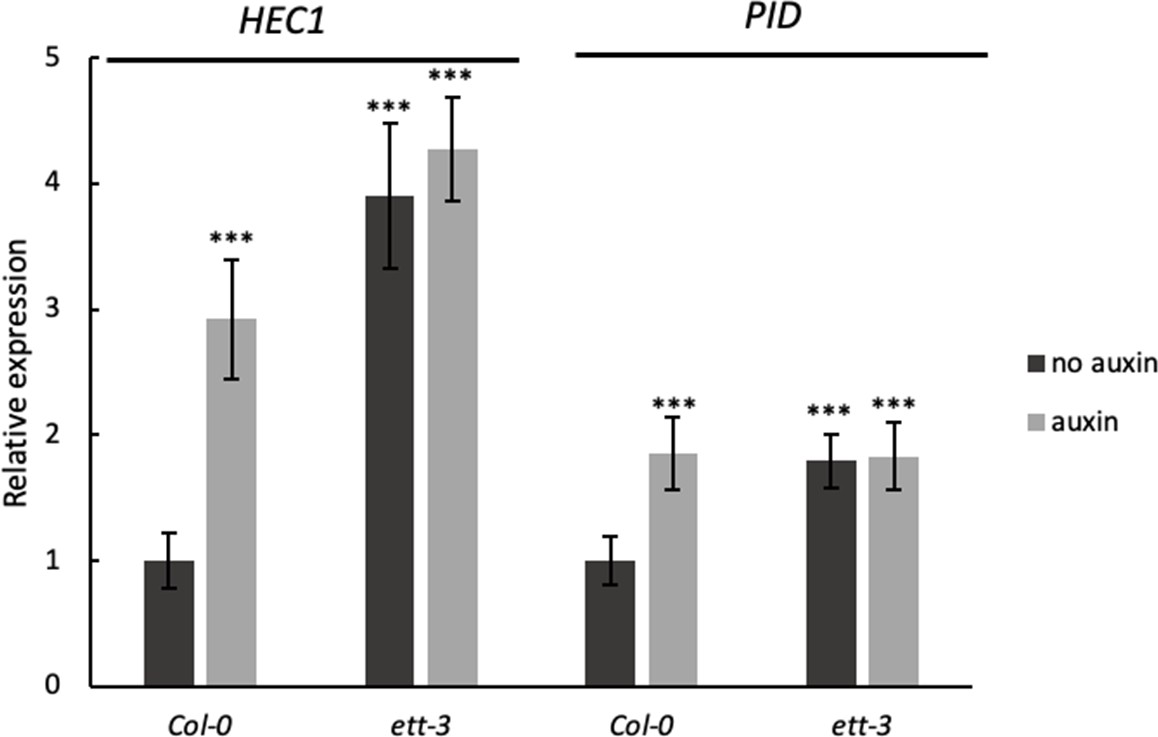
Expression of HEC1 and PID in Col-0 and ett-3.
In wild-type gynoecia HEC1 and PID are up-regulated upon auxin treatment while both genes are constitutively up-regulated in ett-3. Treatment with 100 µM IAA does not affect HEC1 and PID expression in the ett-3 mutant suggesting that ETT acts as a transcriptional repressor. Asterisks indicate significant change upon auxin treatment compared to untreated Col-0 (*** indicating p<0.0001). Shown are mean ± standard deviation of three biological replicates. See Figure 2—source data 2 for statistical analyses.
Figure 3 with 3 supplements
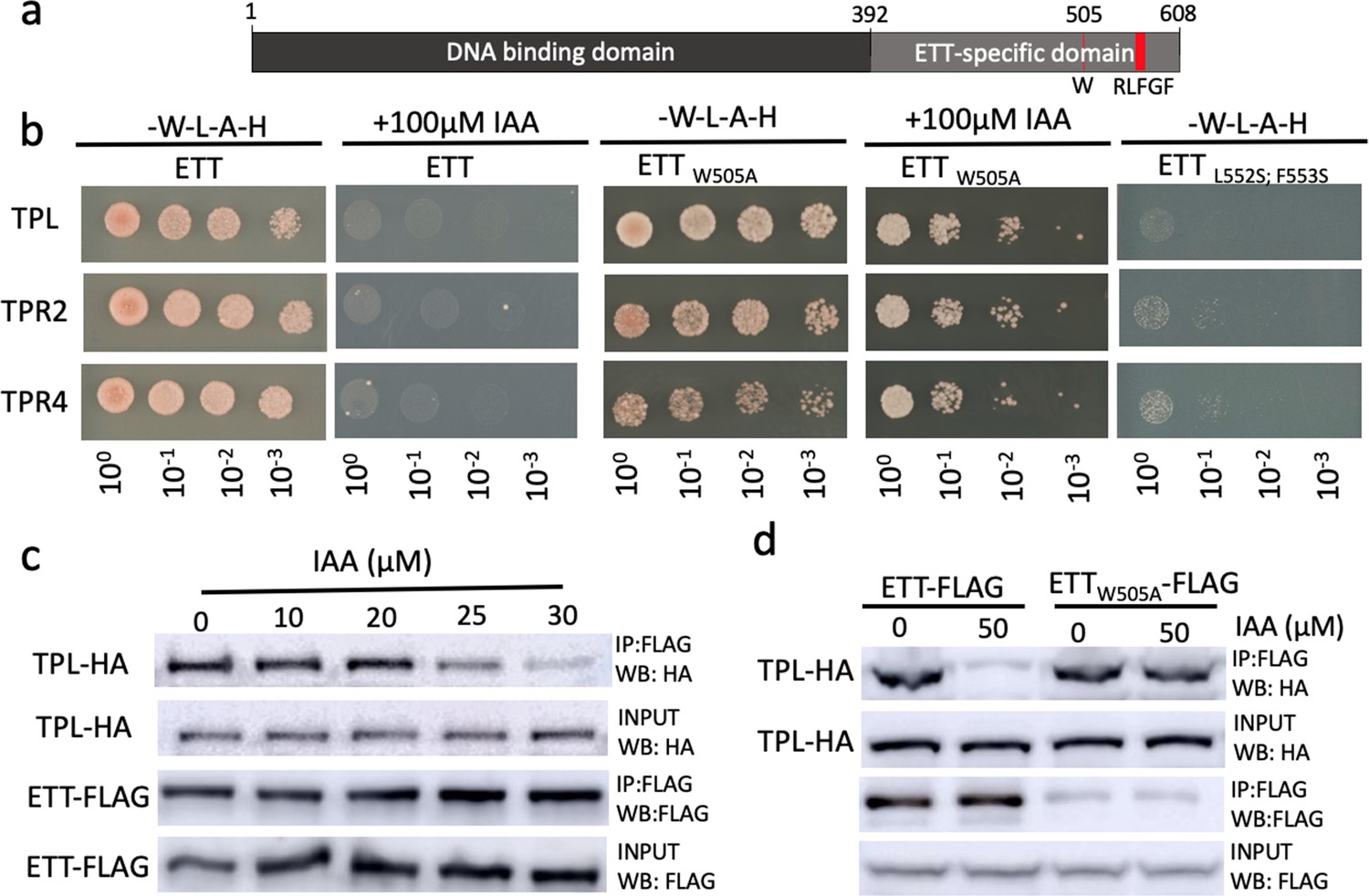
ETT interacts with members of the TPL/TPR co-repressor family in an auxin-sensitive manner.
(a) Schematic representation of full-length ETT protein highlighting an EAR-like motif and tryptophan in position 505 (W) in the C-terminal ETT-specific domain. DBD, DNA-binding domain. (b) Y2H showing that ETT interacts with TPL, TPR2 and TPR4. These interactions depend on the identified C-terminal RLFGF motif and are auxin-sensitive. DBD, DNA-binding domain. (c) Co-IP revealing that ETT interacts with TPL in an auxin-sensitive manner with increasing IAA concentrations weakening the interaction. (d) Co-IP showing that mutating the tryptophan in position 505 of ETT (ETTW505A) leads to loss of auxin sensitivity in the interaction between ETT and TPL.
-
Figure 3—source data 1
Output of statistical tests.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig3-data1-v2.xlsx
Figure 3—figure supplement 1
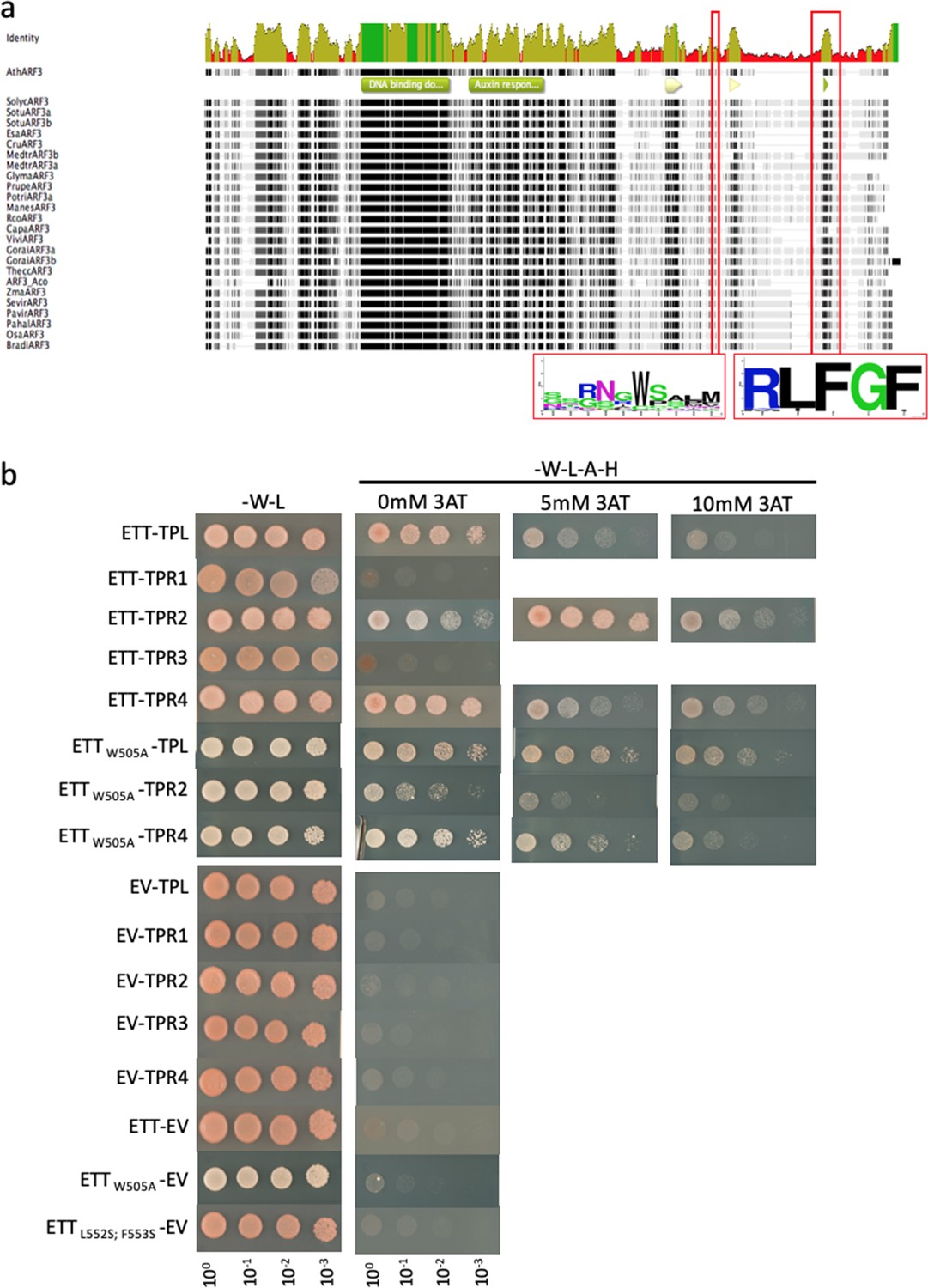
ETT can interact with several members of the TPL/TPR co-repressor family through a conserved EAR-like motif.
(a) Alignment of ETT protein sequences of 22 species identified a conserved repressive motif (RLFGF) at its C-terminal domain. A logo of the sequence surrounding the conserved W505 is also shown. (b) ETT interacts with several members of the TPL/TPR co-repressor family in Y2H. Additionally, controls for Figure 2 are shown.
Figure 3—figure supplement 2
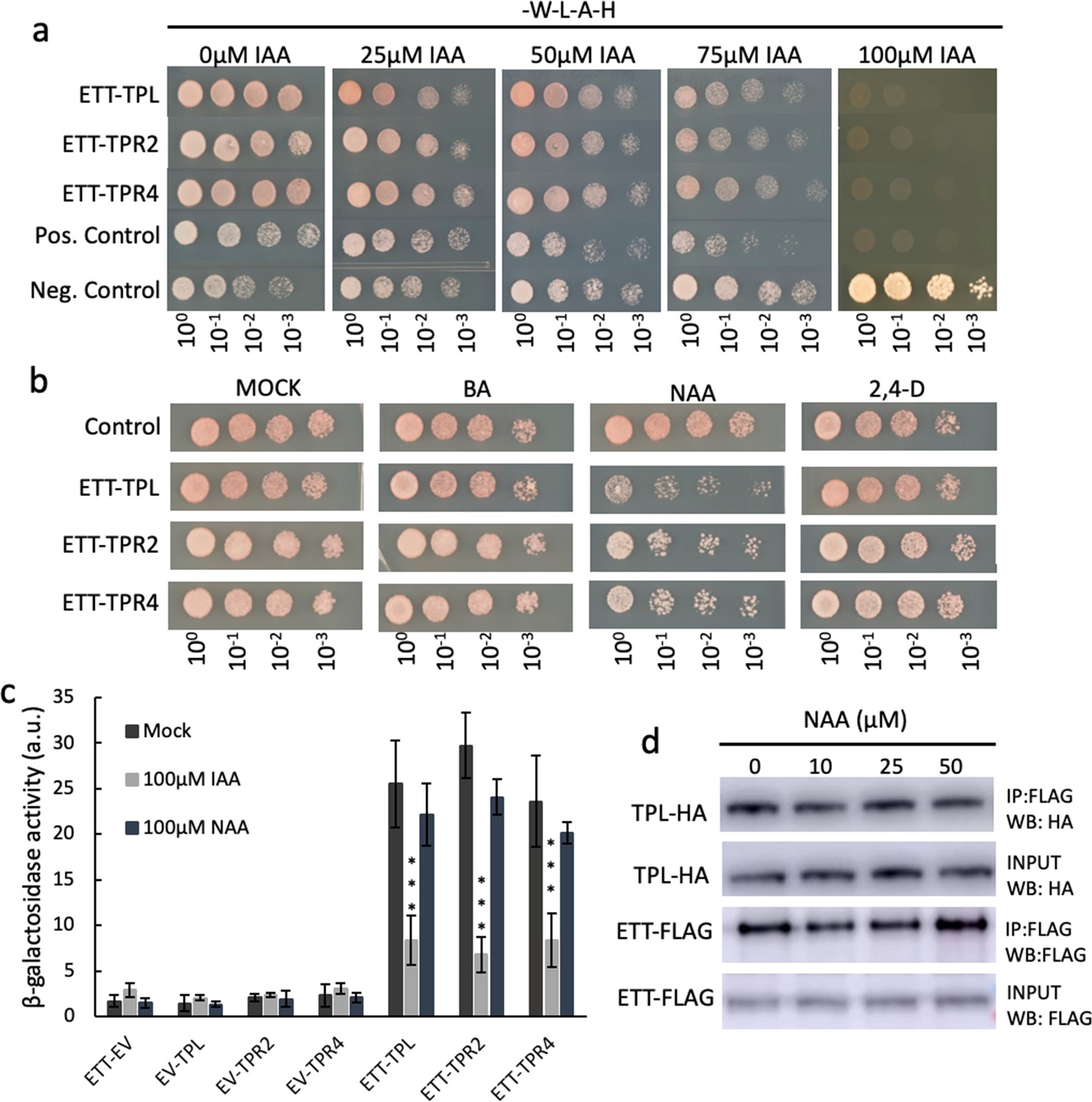
Interaction between ETT and TPL, TPR2 and TPR4 is auxin-sensitive and specific to IAA.
(a) In Y2H increasing concentrations of IAA lead to reduction of yeast growth abolishing the interaction between ETT and its partners. The interactions are, therefore, auxin-sensitive. (b) Y2H to test specificity of auxin-sensitivity using benzoic acid (BA), NAA, and 2,4D in a yeast growth assay. The data suggest that the auxin-sensitivity observed in (a) is IAA-specific. (c) Y2H-based ONPG assay quantifying β-galactosidase activity as a measure of interaction strength.( d) Co-IP experiments show that the interaction between ETT and TPL cannot be disrupted by the synthetic auxin, NAA. The data support that the ETT TPL/TPR interactions are sensitive to IAA but not to NAA. ***p<0.0001; Shown are mean ± standard deviation of three biological replicates. See Figure 3—source data 1 for statistical analyses.
Figure 3—figure supplement 3

Original western blot images for Co-immunoprecipitation assays.
The red boxes indicate the regions used in Figure 3c,d and Figure 3—figure supplement 2d.
Figure 4 with 1 supplement
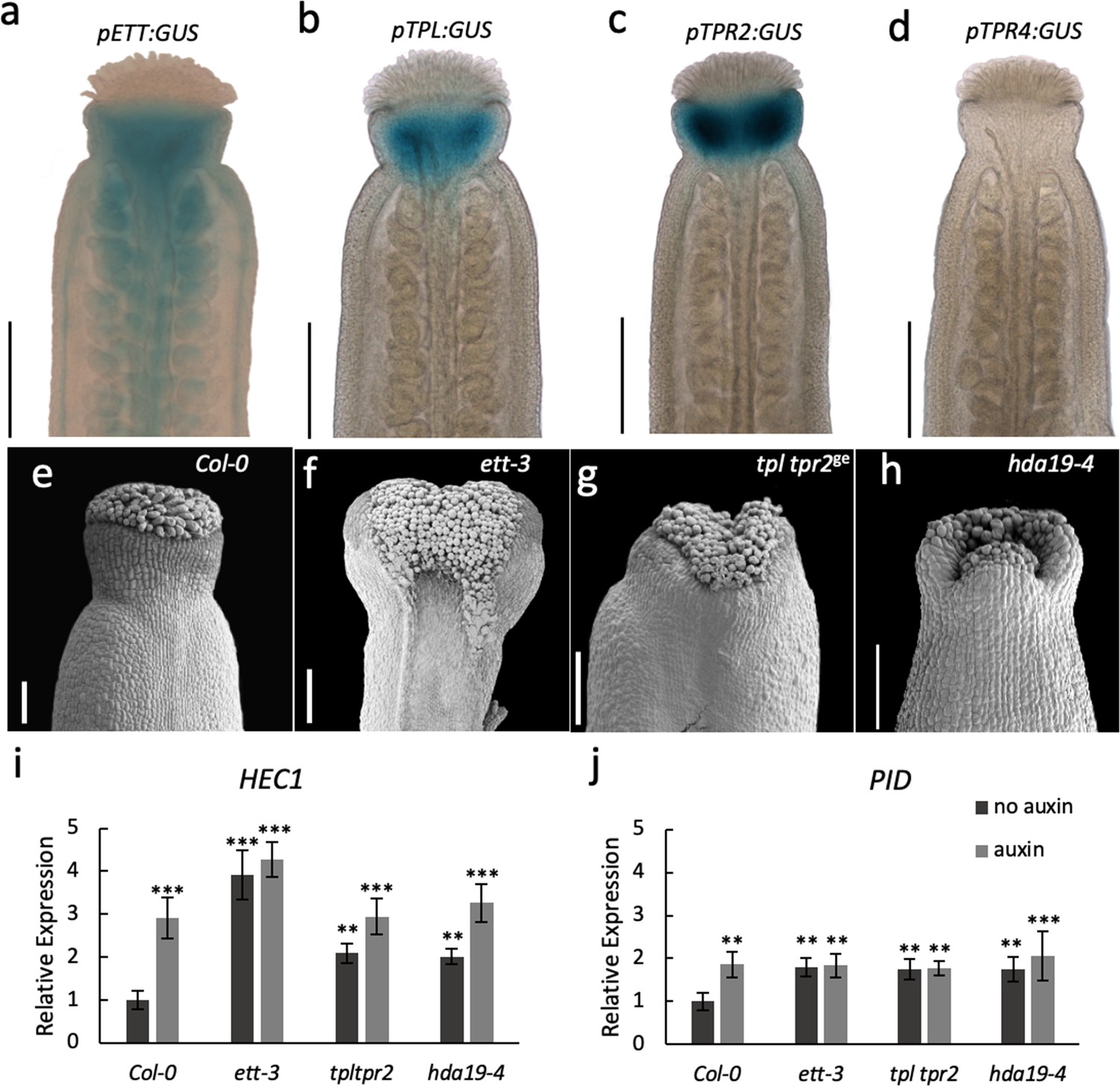
ETT, TPL/TPR2 and HDA19 co-operatively regulate gene expression to facilitate gynoecium development.
(a–d) Promoter GUS expression analysis of pETT:GUS (a), pTPL:GUS (b), pTPR2:GUS (c) and pTPR4:GUS (d) revealed that ETT, TPL and TPR2 but not TPR4 are co-expressed in the Arabidopsis style. Scale bar = 300 μm. (e–h) Gynoecium phenotypes of wild-type (e), ett-3 (f) tpl tpr2ge (g) and hda19-4 (h). Scale bar = 100 μm. (i j,) HEC1 (i) and PID (j) are constitutively mis-regulated in ett-3, tpl tpr2ge and hda19-4 gynoecia. This misregulation is unaffected by treatment with 100 µM IAA. ***p-values<0.0001; Shown are mean ± standard deviation of three biological replicates. Differences between untreated and IAA-treated mutants are not significant. See Figure 4—source data 1 for statistical analyses.
-
Figure 4—source data 1
Output of statistical tests.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig4-data1-v2.xlsx
Figure 4—figure supplement 1
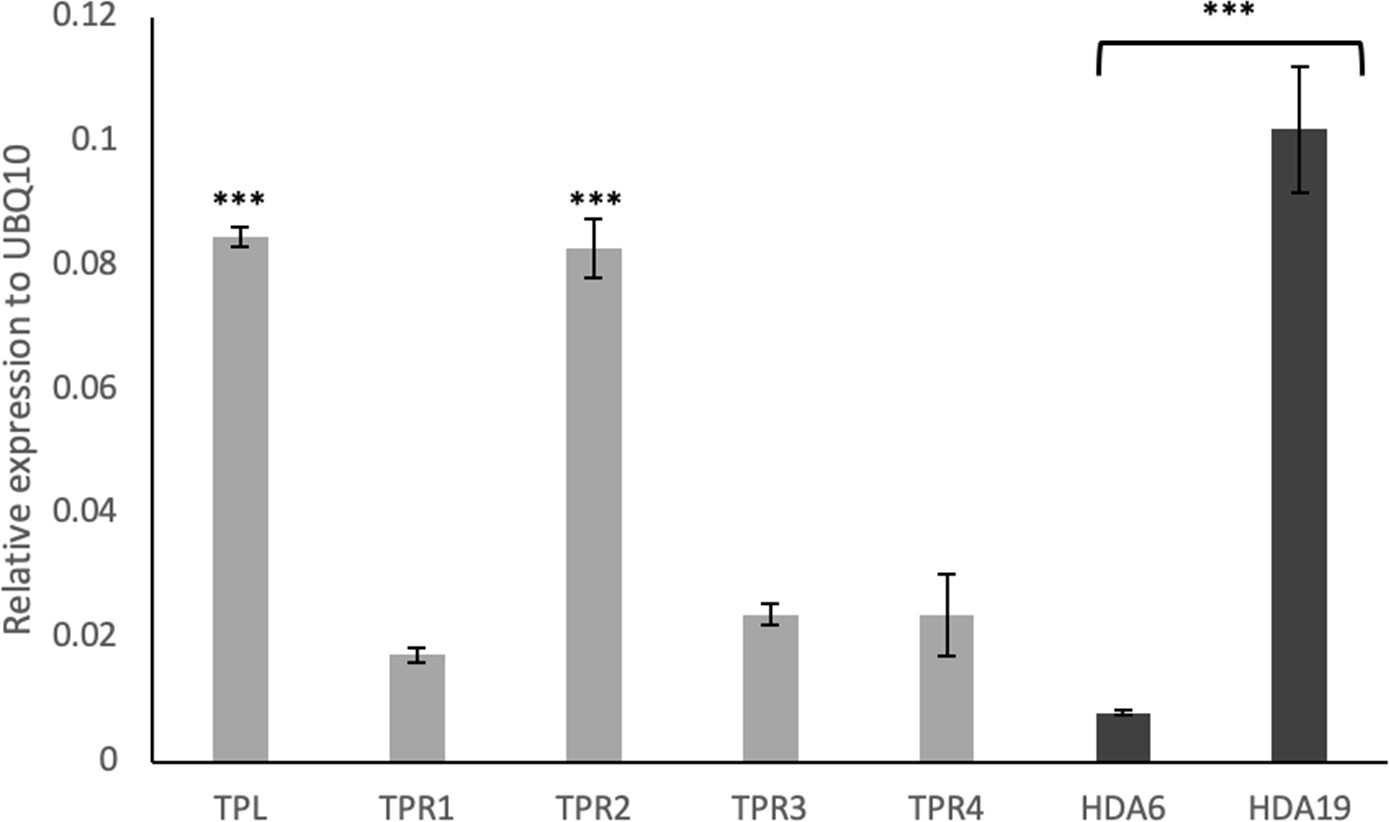
Expression of TPL, TPRs and HDAs genes in the gynoecium.
Expression analysis using qRT-PCR in wild-type gynoecia showed that TPL and TPR2 are more strongly expressed than TPR1,3 and 4. Likewise, HDA19 exhibits higher expression compared to HDA6. ***p-values<0.0001; Shown are mean ± standard deviation of three biological replicates. See Figure 4—source data 1 for statistical analyses.
Figure 5

ETT, TPL and HDA19 co-operatively regulate HEC1 and PID by modulating chromatin acetylation.
(a–c) Chromatin immunoprecipitation (ChIP) shows ETT (a), TPL (b) and HDA19 (c) binding to conserved regions of HEC1 and PID loci in the absence of IAA (dark grey columns). In the presence of IAA (light grey columns), ETT association is unchanged, while both TPL and HDA19 interactions are reduced. A WUS gene fragment served as negative control (NC). (d e,) H3K27ac accumulation (from ChIP analysis) along the HEC1 (d) and PID (e) loci in wild-type (Col-0) and ett-3 plants ± treatment with 100 µM IAA. Numbers on the x axes are distances to the Transcription Start Site (TSS). The schematic of the loci is shown below each panel. Dashed boxes represent ETT binding regions. (f) Qualitative schematic illustration of alternative TIR1/AFB independent auxin signaling. Under low auxin conditions an ETT-TPL-HDA19 complex binds to ETT-target genes keeping their chromatin environments repressed, through de-acetylation. High nuclear auxin concentrations abolish the ETT-TPL-HDA19 complex through direct ETT-auxin interaction. This leads to an accumulation of histone acetylation and up-regulation of ETT-target genes. Values in a–e) are means ± standard deviation of three biological replicates. See Figure 5—source data 1 for statistical analyses.
-
Figure 5—source data 1
Output of statistical tests.
- https://cdn.elifesciences.org/articles/51787/elife-51787-fig5-data1-v2.xlsx
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information | ||||
|---|---|---|---|---|---|---|---|---|
| Gene (Arabidopsis thaliana) | ETTIN (ETT) | The Arabidopsis Information Resource | AT2G33860 | |||||
| Gene (Arabidopsis thaliana) | TOPLESS (TPL) | The Arabidopsis Information Resource | AT1G15750 | |||||
| Gene (Arabidopsis thaliana) | TOPLESS- RELATED 1 (TPR1) | The Arabidopsis Information Resource | AT1G80490 | |||||
| Gene (Arabidopsis thaliana) | TOPLESS-RELATED 2 (TPR2) | The Arabidopsis Information Resource | AT3G16830 | |||||
| Gene (Arabidopsis thaliana) | TOPLESS-RELATED 3 (TPR3) | The Arabidopsis Information Resource | AT5G27030 | |||||
| Gene (Arabidopsis thaliana) | TOPLESS-RELATED 4 (TPR4) | The Arabidopsis Information Resource | AT3G15880 | |||||
| Gene (Arabidopsis thaliana) | HISTONE DEACETYLASE 6 (HDA6) | The Arabidopsis Information Resource | AT5G63110 | |||||
| Gene (Arabidopsis thaliana) | HISTONE DEACETYLASE 19 (HDA19) | The Arabidopsis Information Resource | AT4G38130 | |||||
| Gene (Arabidopsis thaliana) | HECATE 1 (HEC1) | The Arabidopsis Information Resource | AT5G67060 | |||||
| Gene (Arabidopsis thaliana) | PINOID (PID) | The Arabidopsis Information Resource | AT2G34650 | |||||
| Genetic reagent Arabidopsis thaliana | Col-0 | widely distributed | ||||||
| Genetic reagent Arabidopsis thaliana | hda19-4 | Kim et al., 2008 | SALK_13944 | |||||
| Genetic reagent Arabidopsis thaliana | tir1-1 afb2-3 afb3-4 | Parry et al., 2009 | ||||||
| Genetic reagent Arabidopsis thaliana | tpl | European Arabidopsis Stock Centre | SALK_034518C | |||||
| Genetic reagent Arabidopsis thaliana | ett-3 | Simonini et al., 2017; Sessions et al., 1997 | AT2G33860 | |||||
| Genetic reagent Arabidopsis thaliana | pETT:GUS | Ng et al., 2009 | AT2G33860 | |||||
| Genetic reagent Arabidopsis thaliana | pETT:ETT-GFP | Simonini et al., 2016 | AT2G33860 | |||||
| Genetic reagent Arabidopsis thaliana | pTPL:TPL-GFP | Pi et al., 2015 | AT1G15750 | |||||
| Genetic reagent Arabidopsis thaliana | p35S:HDA19-GFP | Pi et al., 2015 | AT4G38130 | |||||
| Genetic reagent Arabidopsis thaliana | pTIR1:ccvTIR1 | Szemenyei et al., 2008 | ||||||
| Genetic reagent Arabidopsis thaliana | tpl tpr2ge | this paper | AT1G15750, AT3G16830 | Further details in the Materials and methods section | ||||
| Genetic reagent Arabidopsis thaliana | pTPL:GUS | this paper | AT1G15750 | Further details in the Materials and methods section | ||||
| Genetic reagent Arabidopsis thaliana | pTPR2:GUS | this paper | AT3G16830 | Further details in the Materials and methods section | ||||
| Genetic reagent Arabidopsis thaliana | pTPR4:GUS | this paper | AT3G15880 | Further details in the Materials and methods section | ||||
| Genetic reagent Arabidopsis thaliana | p35S:CDGVG p6xGAL4UAS:ETT-FLAG | this paper | Further details in the Materials and methods section | |||||
| Antibody | Mouse anti-FLAG | Abcam | ab49763 | monoclonal, conjugated with HRP | ||||
| Antibody | Mouse anti-HA | Abcam | ab173826 | monoclonal, conjugated with HRP | ||||
| Antibody | Mouse anti-GFP | Roche | 11814460001 | monoclonal | ||||
| Antibody | Rabbit anti-H3K27ac | Abcam | ab4729 | polyclonal | ||||
| Antibody | Rabbit anti-H3 | Abcam | ab1791 | polyclonal | ||||
| Recombinant DNA reagent | pGWB14; p35s:HA-TPL | Espinosa-Ruiz et al., 2017 | ||||||
| Recombinant DNA reagent | pICH47732; p35s:ETT-3xFLAG | this paper | Backbone Addgene #48000 | |||||
| Recombinant DNA reagent | pICH47732; p35s: ETTW505A-3xFLAG | this paper | Backbone Addgene #48000 | |||||
| Recombinant DNA reagent | pGADT7; ETT | this paper | Further details in the Materials and methods section | |||||
| Recombinant DNA reagent | pGADT7; ETTW505A | this paper | Further details in the Materials and methods section | |||||
| Recombinant DNA reagent | pGADT7; ETTL552S; F553S | this paper | Further details in the Materials and methods section | |||||
| Recombinant DNA reagent | pGBKT7; TPL | Causier et al., 2012 | ||||||
| Recombinant DNA reagent | pGBKT7; TPR1 | Causier et al., 2012 | ||||||
| Recombinant DNA reagent | pGBKT7; TPR2 | Causier et al., 2012 | ||||||
| Recombinant DNA reagent | pGBKT7; TPR3 | Causier et al., 2012 | ||||||
| Recombinant DNA reagent | pGBKT7; TPR4 | Causier et al., 2012 | ||||||
| Recombinant DNA reagent | pESPRIT; ES388-594 | Simonini et al., 2018 | ||||||
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Direct ETTIN-auxin interaction controls chromatin states in gynoecium development
eLife 9:e51787.
https://doi.org/10.7554/eLife.51787




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}