fruitless tunes functional flexibility of courtship circuitry during development
- The Key Laboratory of Developmental Genes and Human Disease, School of Life Science and Technology, Southeast University, China
- Co-innovation Center of Neuroregeneration, Nantong University, China
Abstract
Drosophila male courtship is controlled by the male-specific products of the fruitless (fruM) gene and its expressing neuronal circuitry. fruM is considered a master gene that controls all aspects of male courtship. By temporally and spatially manipulating fruM expression, we found that fruM is required during a critical developmental period for innate courtship toward females, while its function during adulthood is involved in inhibiting male–male courtship. By altering or eliminating fruM expression, we generated males that are innately heterosexual, homosexual, bisexual, or without innate courtship but could acquire such behavior in an experience-dependent manner. These findings show that fruM is not absolutely necessary for courtship but is critical during development to build a sex circuitry with reduced flexibility and enhanced efficiency, and provide a new view about how fruM tunes functional flexibility of a sex circuitry instead of switching on its function as conventionally viewed.
eLife digest
Innate behaviors are behaviors that do not need to be learned. They include activities such as nest building in birds and web spinning in spiders. Another behavior that has been extensively studied, and which is generally considered to be innate, is courtship in fruit flies. Male fruit flies serenade potential mates by vibrating their wings to create a complex melody. This behavior is under the control of a gene called ‘fruitless’, which gives rise to several distinct proteins, including one that is unique to males. For many years, this protein – called FruM – was thought to be the master switch that activates courtship behavior.
But recent findings have challenged this idea. They show that although male flies that lack FruM fail to show courtship behaviors if raised in isolation, they can still learn them if raised in groups. This suggests that the role of FruM is more complex than previously thought. To determine how FruM controls courtship behavior, Chen et al. have used genetic tools to manipulate FruM activity in male flies at different stages of the life cycle and distinct cells of the nervous system.
The results revealed that FruM must be present during a critical period of development – but not adulthood – for male flies to court females. However, FruM strongly influences the type of courtship behavior the male flies display. The amount and location of FruM determines whether males show heterosexual, homosexual or bisexual courtship behaviors. Adult flies with lower levels of FruM show an increase in homosexual courtship and a decrease in heterosexual courtship.
These findings provide a fresh view on how a master gene can generate complex and flexible behaviors. They show that fruitless, and the FruM protein it encodes, work distinctly at different life cycles to modify the type of courtship behavior shown by male flies, rather than simply switching courtship behavior on and off. Exactly how FruM acts within the fruit fly brain to achieve these complex effects requires further investigation.
Introduction
Drosophila male courtship is one of the best understood innate behaviors in terms of genetic and neuronal mechanisms (Dickson, 2008; Yamamoto and Koganezawa, 2013). It has been well established that the fruitless (fru) gene and its expressing neurons control most aspects of such innate behavior (Ito et al., 1996; Manoli et al., 2005; Ryner et al., 1996; Stockinger et al., 2005). The male-specific products of the P1 promoter of the fru gene (fruM) are expressed in ~2000 neurons, which are inter-connected to form a sex circuitry from sensory neurons to motor neurons (Cachero et al., 2010; Lee et al., 2000; Manoli et al., 2005; Stockinger et al., 2005; Usui-Aoki et al., 2000; Yu et al., 2010). fruM function is necessary for the innate courtship behavior and sufficient for at least some aspects of courtship (Baker et al., 2001; Demir and Dickson, 2005; Manoli et al., 2005). Thus, the study of fruM function in controlling male courtship serves as an ideal model to understand how innate complex behaviors are built into the nervous system by regulatory genes (Baker et al., 2001).
Although fruM serves as a master gene controlling Drosophila male courtship, we recently found that males without fruM function, although did not court if raised in isolation, were able to acquire at least some courtship behaviors if raised in groups (Pan and Baker, 2014). Such fruM-independent but experience-dependent courtship acquisition requires another gene in the sex determination pathway, the doublesex (dsx) gene (Pan and Baker, 2014). dsx encodes male- and female-specific DSX proteins (DSXM and DSXF, respectively) (Burtis and Baker, 1989), and DSXM is expressed in ~700 neurons in the central nervous system (CNS), the majority of which also express fruM (Rideout et al., 2010; Robinett et al., 2010). It has been found that the fruM and dsxM co-expressing neurons are required for courtship in the absence of fruM function (Pan and Baker, 2014). Thus fruM-expressing neurons, especially those co-expressing dsxM, control the expression of courtship behaviors even in the absence of FRUM function. Indeed, although the gross neuroanatomical features of the fruM-expressing circuitry are largely unaffected by the loss of fruM (Manoli et al., 2005; Stockinger et al., 2005), detailed analysis revealed morphological changes of many fruM-expressing neurons (Cachero et al., 2010; Kimura et al., 2005; Kimura et al., 2008; Mellert et al., 2010). Recent studies further reveal that FRUM specifies neuronal development by recruiting chromatin factors and changing chromatin states, and also by turning on and off the activity of the transcription repressor complex (Ito et al., 2012; Ito et al., 2016; Sato et al., 2019a; Sato et al., 2019b; Sato and Yamamoto, 2020).
That FRUM functions as a transcription factor to specify development and/or physiological roles of certain fruM-expressing neurons, and perhaps the interconnection of different fruM-expressing neurons to form a sex circuitry raises important questions regarding when fruM functions and how it contributes to the sex circuitry (e.g., how the sex circuitry functions differently with different levels of FRUM), especially in the background that fruM is not absolutely necessary for male courtship (Pan and Baker, 2014). To at least partially answer these questions, we temporally or spatially knocked down fruM expression and compared courtship behavior in these males with that in wild-type males or fruM null males and revealed crucial roles of fruM during a narrow developmental window for the innate courtship toward females. We also found that the sex circuitry with different fruM expression has distinct function such that males could be innately heterosexual, homosexual, bisexual, or without innate courtship but could acquire such behavior in an experience-dependent manner. Thus, fruM tunes functional flexibility of the sex circuitry instead of switching on its function as conventionally viewed.
Results
fruM is required during pupation for regular neuronal development and female-directed courtship
To specifically knockdown fruM expression, we used a microRNA targeting fruM (UAS-fruMi at attp2 or attp40) and a scrambled version as a control (UAS-fruMiScr at attp2) as previously used (Chen et al., 2017; Meissner et al., 2016). Driving the fruM microRNA by fruGAL4 specifically knocked down mRNA of fruM, but not the common form of fru (Figure 1—figure supplement 1A–C). We firstly tested male courtship without food in the behavioral chamber. Knocking down fruM in all the fruGAL4-labeled neurons eliminated male courtship toward females (courtship index [CI], which is the percentage of observational time that males displayed courtship, is nearly 0) (Figure 1A), consistent with previous findings that fruM is required for innate male–female courtship (Demir and Dickson, 2005; Pan and Baker, 2014). As fruGAL4 drives expression throughout development and adulthood (Figure 1—figure supplement 1D–K), we set out to use a temperature-dependent tub-GAL80ts transgene to restrict UAS-fruMi expression (e.g., at 30°C) at different developmental stages. We raised tub-GAL80 ts/+; fruGAL4/UAS-fruMi flies at 18°C (permissive for GAL80ts that inhibits GAL4 activity) and transferred these flies to fresh food vials every 2 days. In this way, we generated tub-GAL80 ts/+; fruGAL4/UAS-fruMi flies at nine different stages from embryos to adults and incubated all flies at 30°C to allow fruM knockdown for 2 days, then placed all flies back to 18°C until courtship test (Figure 1B). We found that males with fruM knocked down at stage 5 for 2 days, matching the pupation phase, rarely courted (CI < 10%), and none successfully mated, while males with fruM knocked down near this period (stages 4 and 6) showed a partial courtship or mating deficit, and males with fruM knocked down at earlier or later stages showed strong courtship toward females and successful mating (Figure 1C,D).
Figure 1 with 4 supplements see all
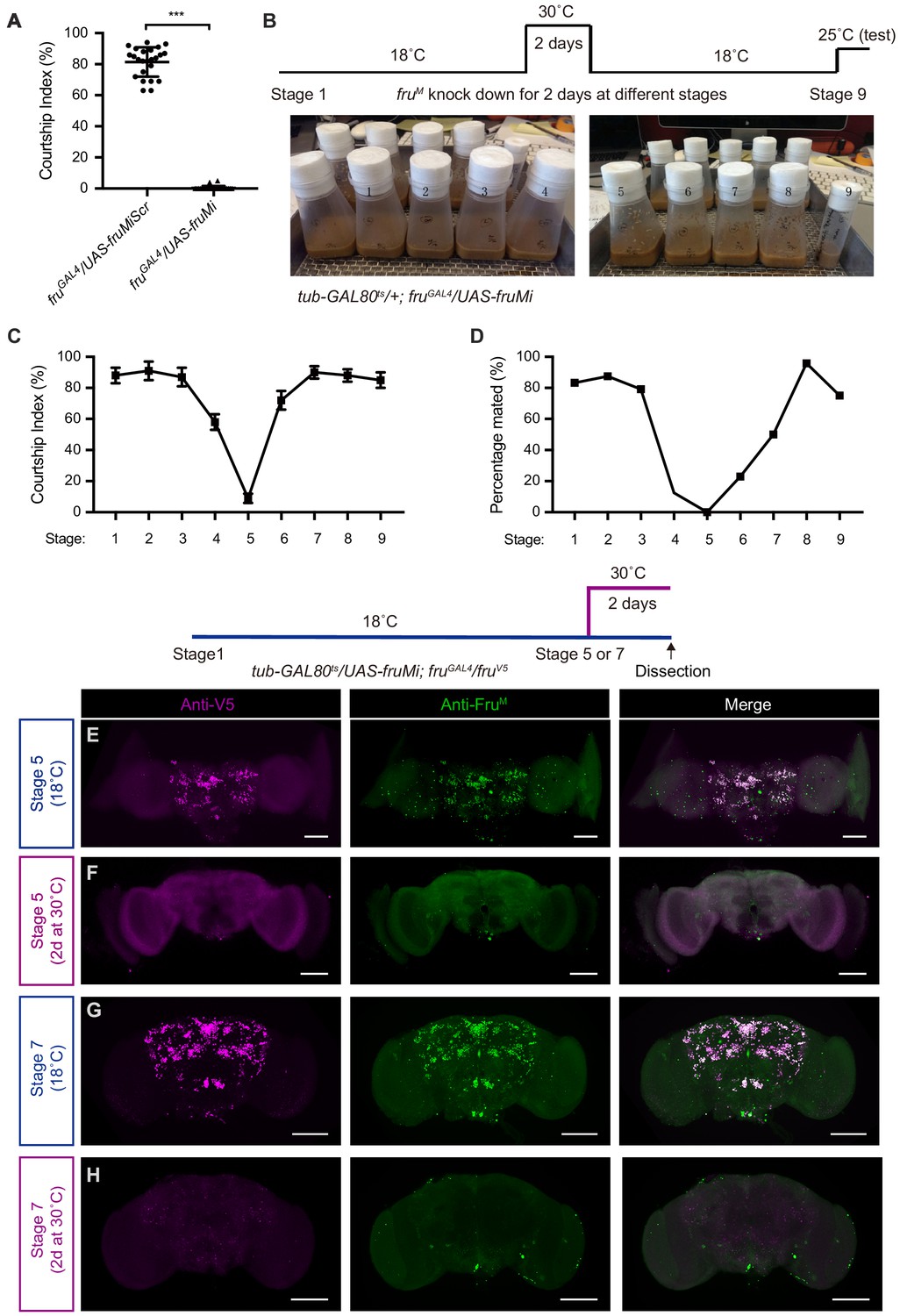
fruM is required during pupation for female-directed courtship in adult males.
(A) Knocking down fruM using RNAi throughout development and adulthood eliminated male courtship toward virgin females. n = 24 for each. ***p<0.001, unpaired t-test. (B) A schematic of genetic strategy to knockdown fruM at different developmental stages for 2 days. Stages 1–9 refer to specific developmental stages from embryos to newly eclosed adults with interval of 2 days. (C and D) Courtship indices of males with fruM knocked down at specific developmental stages as indicated above toward virgin females. Males with fruM knocked down at stage 5 for 2 days (a period of pupation from stage 5 to 6, see above picture) rarely courted virgin females (C), and none successfully mated (D). Knocking down fruM at stages near 5 (e.g., stage 4 or 6) also partially impairs courtship and mating success. Knocking down fruM at earlier or later stages has no obvious effect on courtship and mating. n = 24 for each. Error bars indicate SEM. (E–H) Two day heat shock at 30°C effectively knocks down fruM expression during development. Anti-V5 and anti-FruM signals are dramatically decreased after heat shock at stage 5 (E and F) or 7 (G and H) in tub-GAL80ts/UAS-fruMi; fruGAL4/fruV5 males. Scale bars, 100 μm. Representative of five samples each.
-
Figure 1—source data 1
Source data for Figure 1.
- https://cdn.elifesciences.org/articles/59224/elife-59224-fig1-data1-v2.xlsx
To validate efficiency of fruM knockdown during specific developmental periods, we generated an antibody against FruM as well as a V5 knock-in into the fru gene (fruV5) to visualize FruM expression. Both tools successfully labeled male-specific FruM proteins (Figure 1—figure supplement 2), and there is almost perfect overlap of the two markers (Figure 1E,G). Note that the anti-FruM antibody also labeled several pairs of false-positive neurons in both wild-type and fruM mutants (Figure 1—figure supplement 2), indicating the strong but not perfect specificity of this antibody (Figure 1—figure supplement 2B–D). To test whether 2 day heat shock at 30°C is sufficient to knockdown fruM expression, we dissected brains of tub-GAL80ts/UAS-fruMi; fruGAL4/fruV5 males immediately after 2 day heat shock at stage 5 or 7 and found that anti-V5 and anti-FruM signals were both dramatically decreased, such that only a small fraction of neurons could be weakly labeled; in contrast, control males with the same age but raised at 18°C have regular anti-V5 and anti-FruM signals (Figure 1E–H). These results indicate that induction of fruM microRNA during development for 2 days could effectively knockdown fruM expression.
As induced fruM microRNA may not be degraded immediately and has longer effect, we further tested to how much extent such knockdown effect may last. Thus, we dissected brains of adult tub-GAL80ts/UAS-fruMi; fruGAL4/fruV5 males that have been heat shocked for 2 days at different developmental stages (from stages 1 to 9) and found that males that have been heat shocked at earlier stages (from stages 1 to 5) still have strong FruM expression (Figure 1—figure supplement 3A–F), suggesting effective restore of FruM expression after transferring at 18°C. However, males that have been heat shocked at later stages (stages 6–9) have obviously reduced FruM expression (Figure 1—figure supplement 3G–J), suggesting a partial restore of FruM expression, probably due to prolonged fruM microRNA effect. Note that knocking down fruM expression at these later stages has partial (stage 6) or no effect (other stages) on male courtship, comparing with fruM knockdown at stage 5 that almost eliminated male courtship. Together these results indicate a critical developmental period during pupation (from late larvae at stage 5 to early pupas at stage 6) where fruM is required for adult male courtship toward females.
We reasoned that fruM function during pupation may be involved in neuronal development for circuit construction. Thus we set out to examine the morphology of a subset of fruM-positive gustatory receptor neurons (GRNs) innervating the ventral nerve cord (VNC) in tub-GAL80ts/UAS-mCD8GFP; fruGAL4/UAS-fruMi males that have been heat shocked for 2 days in different developmental stages, as it has been found that fruM is required for the male-specific midline crossing of these GRNs (Mellert et al., 2010). We found that these GRNs were only labeled in males that have been heat shocked after stage 4, probably because these GRNs were developed after stage 4 (Figure 1—figure supplement 4A–C), consistent with a previous study (Mellert et al., 2012). Interestingly, we found that all males heat shocked at stage 5 for 2 days showed defect of midline crossing in these GRNs, and 60% of males heat shocked at stage 6 for 2 days showed defect of midline crossing, while all males heat shocked after stage 6 showed regular midline crossing (Figure 1—figure supplement 4C,D). Males heat shocked for 4 days during adulthood also have regular midline crossing (Figure 1—figure supplement 4C,D). These results clearly showed a critical developmental period during pupation where FruM functions to ensure regular development of GRNs and enable innate male courtship toward females.
fruM function during adulthood inhibits male–male courtship
As knocking down fruM at stage 9 when flies were newly eclosed did not affect male courtship (CI > 80%) and mating success (Figure 1C,D), we further tested the role of fruM in adulthood using different approaches. We set out to express the female-specific transformer (traF) gene (Baker and Ridge, 1980; McKeown et al., 1988) to feminize all fruGAL4 labeled neurons, in addition to the above fruM RNAi experiments. We express UAS-traF or UAS-fruMi in all the fruGAL4-labeled neurons specifically during adulthood for 4 days before test (see procedure above each figure) for single-pair male–female, male–male, and male chaining (in groups of eight males) behaviors. We found that overexpression of traF in all fruGAL4 labeled neurons during adulthood for 4 days did not affect male–female courtship (Figure 2A), but slightly increased male–male (Figure 2B) and male chaining behaviors (Figure 2C). Furthermore, knocking down fruM in all fruGAL4-labeled neurons during adulthood for 4 days did not affect male–female (Figure 2A) or male–male courtship (Figure 2B), but significantly increased male chaining behaviors (Figure 2C). We also checked FruM expression in males that have been heat shocked for 4 days during adulthood using anti-V5 and anti-FruM antibodies, and found that FruM expression was almost eliminated, while control males have regular FruM expression (Figure 2D,E). These results indicate that although fruM function during adulthood is dispensable for female-directed courtship, it is involved in inhibiting male–male courtship behaviors. Thus, FruM has distinct functions during development and adulthood for male courtship behaviors.
Figure 2

fruM functions during adulthood to inhibit male–male courtship behaviors.
(A–C) Courtship behaviors performed by males that express traF or fruMi specifically during adulthood for 4 days. For male–female courtship (A), n = 17, 26, 23, 23, 24, 27, 24, and 28, respectively (from left to right), n.s., not significant, unpaired t-test. For single-pair male–male courtship (B), n = 18 for each. n.s., not significant, *p<0.05, unpaired t-test. For male chaining among eight males as a group (C), n = 8, 8, 8, 10, 8, 18, 8, and 18, respectively (from left to right). n.s., not significant, *p<0.05, **p<0.01, ***p<0.001, Mann–Whitney U test. Error bars indicate SEM. Genotypes as indicated. (D and E) Anti-V5 and anti-FruM signals are dramatically decreased after heat shock during adulthood for 4 days in tub-GAL80ts/UAS-fruMi; fruGAL4/fruV5 males. Scale bars, 100 μm. Representative of five samples each.
-
Figure 2—source data 1
Source data for Figure 2.
- https://cdn.elifesciences.org/articles/59224/elife-59224-fig2-data1-v2.xlsx
fruM expression determines courtship modes
The above results indicate crucial roles of fruM during pupation for female-directed courtship in adult males. We reasoned that fruM function during pupation may specify the construction of courtship circuitry and affects female-directed courtship as well as other courtship behaviors, especially given our previous findings that fruM null males were able to acquire courtship behavior after group-housing (Pan and Baker, 2014). Thus, we set out to compare courtship behaviors in males with distinct fruM expression modes, such as with wild-type fruM, systemic low level of fruM, spatially low level of fruM, or completely without fruM function. We tested one-time single-pair male–female and male–male courtship (single housed before test) as well as male chaining in groups of eight males over 3 days on food for better comparison of these courtship assays, as courtship by fruM null males largely depends on food presence (Pan and Baker, 2014). We found that male–male courtship in fruM knocked down males is higher if tested on food, consistent with a courtship promoting role by food (Grosjean et al., 2011; Pan and Baker, 2014), while courtship in wild-type males on food or without food is not changed in our assays (Figure 3—figure supplement 1). We found that wild-type males performed intensive courtship behavior toward virgin females (CI > 80%) and rarely courted males (CI ~0) (Figure 3A). Furthermore, these control males did not show any chaining behavior after grouping from 3 hr to 3 days (ChI = 0) (Figure 3B). In striking contrast, fruM null mutant males rarely courted either females or males (Figure 3C, Figure 3—figure supplement 2A, C, and E); however, these males developed intensive chaining behavior after grouping for 1–3 days (Figure 3D, Figure 3—figure supplement 2B, D, and F). These observations replicated previous findings that there exists a fruM-independent experience and dsxM-dependent courtship pathway (Pan and Baker, 2014; Figure 3E). To compare behavioral differences by fruM null males and fruM RNAi knocked down males that have systemic low level of fruM, we firstly quantified to how much extent the microRNA against fruM (UAS-fruMi at attp40) worked. We found that the fruM mRNA level was reduced to ~40% of that in control males (Figure 3F). Interestingly, while males with fruM knocked down in all fruM neurons rarely courted females (CI ~5%, Figure 3G), they displayed a high level of male–male courtship behavior (CI > 50%, Figure 3G) and constantly high level of male chaining (Figure 3H), dramatically different from fruM null males. These results reveal distinct roles of low fruM (RNAi) and high fruM (wild-type) in regulating male–male and male–female courtship (Figure 3I).
Figure 3 with 4 supplements see all
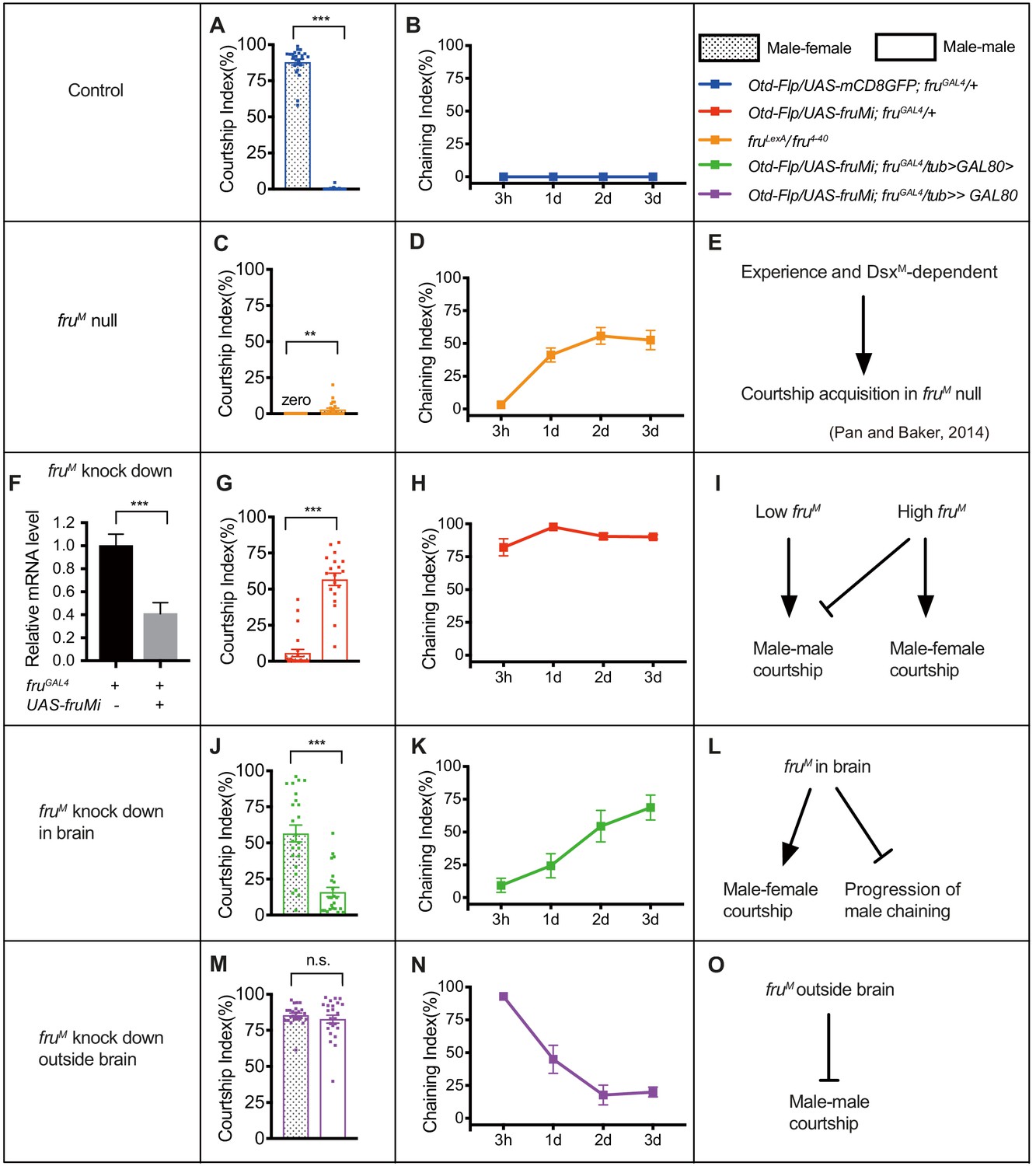
fruM tunes functional flexibility of the fruM circuitry.
(A and B) Wild-type males courted intensively toward virgin females (A, left bar), but rarely courted males (A, right bar) or displayed chaining behavior in groups of eight males (B). n = 24, 24, 8, respectively. ***p<0.001, unpaired t-test. (C) FruLexA/fru4-40 (fruM null) males rarely courted either females or males. n = 24 for each, **p<0.01, Mann–Whitney U test. (D) FruLexA/fru4-40 males did not show chaining behavior after 3 hr group-housing, but developed intensive chaining behavior after1-3 days. n = 8. (E) A summary of courtship acquisition independent of fruM. (F) RNAi against fruM efficiently decreased but not fully eliminated fruM expression. n = 4. ***p<0.001, Mann–Whitney U test. (G) Knocking down fruM in all fruGAL4 neurons generated males that have reversed sexual orientation such that they rarely courted females but intensively courted males. n = 24 and 19, respectively. ***p<0.001, unpaired t-test. (H) Males with fruM knocked down in all fruGAL4 neurons showed intensive chaining behavior at all time points (from 3 hr to 3 days upon group-housing). n = 7. (I) Distinct roles of low fruM (RNAi) and high fruM (wild-type) in regulating male–male and male–female courtship. (J) Males with fruM knocked down in fruGAL4 neurons in the brain had a lower level of courtship toward females, but their sexual orientation was not changed. n = 24 and 23, respectively. ***p<0.001, unpaired t-test. (K) Males with fruM knocked down in fruGAL4 neurons in brain showed low male chaining behavior initially but increasing levels of chaining behavior over 1–3 days. n = 6. (L) A summary of the role of fruM in brain in promoting male–female courtship and suppressing the experience-dependent acquisition or progression of male chaining behavior. (M) Males with fruM knocked down in fruGAL4 neurons outside brain generated bisexual males that have intensive male–female and male–male courtship. n = 24 for each. n.s., not significant, unpaired t-test. (N) Males with fruM knocked down in fruGAL4 neurons outside brain showed high male chaining behavior initially, but decreased levels of chaining behavior over 1–3 days. n = 8. (O) A summary of the role of fruM outside brain in suppressing male–male courtship behavior. Error bars indicate SEM.
-
Figure 3—source data 1
Source data for Figure 3.
- https://cdn.elifesciences.org/articles/59224/elife-59224-fig3-data1-v2.xlsx
To further reveal the role of fruM expression patterns in determining male courtship modes, we tried to spatially knockdown fruM expression using a simple way: fruM in brain and fruM outside brain. We used Otd-Flp expressing FLP specifically in the central brain (Asahina et al., 2014) to divide fruGAL4 expression (Figure 3—figure supplement 3A) into two parts: fruM- and Otd-positive neurons (specifically in brain) in Otd-Flp/UAS-mCD8GFP; fruGAL4/tub>GAL80> males (Figure 3—figure supplement 3B) and fruM-positive but Otd-negative neurons (theoretically outside brain, but still with few in brain) in Otd-Flp/UAS-mCD8GFP; fruGAL4/tub>stop>GAL80 males (Figure 3—figure supplement 3C). We also checked GFP expression in peripheral nervous system in these males and found a few GFP-positive cells in antennae and forelegs in Otd-Flp/UAS-mCD8GFP; fruGAL4/+ males, but rare expression in Otd-Flp/UAS-mCD8GFP; fruGAL4/tub>stop>GAL80 or Otd-Flp/UAS-mCD8GFP; fruGAL4/tub>GAL80> males (Figure 3—figure supplement 3D,E). Thus, we successfully divided fruGAL4 expression into two categories: one with GAL4 expressed in fru+Otd+ neurons in brain and the other with GAL4 expressed in fru+Otd− neurons outside brain. We then used the above intersectional strategy to specifically knockdown fruM expression in or outside brain. To validate such strategy, we used anti-V5 to visualize FruM expression in these males (together with fruV5) and found effective, if not perfect, knockdown of FruM expression spatially (Figure 3—figure supplement 3F–I). We found that males with fruM knocked down specifically in brain had a reduced level of courtship toward females (CI = 56.61 ± 5.86%), but their sexual orientation was not changed as they courted males in a much lower level (CI = 15.94 ± 3.26%, Figure 3J). Furthermore, males with fruM knocked down in brain showed low male chaining behavior initially but increasing levels of chaining behavior over 1–3 days (ChI [3 hr] = 9.35 ± 5.40%, ChI[3d] = 68.82 ± 5.53%, Figure 3K). Knocking down fruM only in a subset of male-specific P1 neurons driven by P1-splitGAL4 in the brain that are important for courtship initiation (Clowney et al., 2015; Kallman et al., 2015; Kimura et al., 2008; Pan et al., 2012; Wu et al., 2019) failed to decrease male–female courtship or induce male chaining behavior (Figure 3—figure supplement 4A,B). These results indicate that fruM function in brain promotes male–female courtship and inhibits acquisition or progression of the experience-dependent chaining behavior (Figure 3L). In contrast, males with fruM knocked down outside brain showed equally intensive male–female and male–male courtship (CI [male–female] = 85.62 ± 1.42%, CI [male–male] = 82.89 ± 2.76%, Figure 3M), indicating an inhibitory role of fruM in these neurons for male–male courtship (Figure 3O). These males performed a high level of male chaining behavior initially (ChI [3 hr] = 92.90 ± 3.08%), but decreased levels of chaining behavior over 1–3 days (ChI [3d] = 20.01 ± 3.75%, Figure 3N), consistent with the above finding that fruM function in the brain which is intact in these males inhibits acquisition or progression of male chaining behavior (Figure 3L). Knocking down fruM in a subset of gustatory receptor neurons expressing ppk23 that respond to female-specific pheromones (Lu et al., 2012; Thistle et al., 2012; Toda et al., 2012) mildly enhanced male–male courtship but did not induce male chaining behavior (Figure 3—figure supplement 4C,D), suggesting a moderate role of fruM in these neurons for inhibiting male–male courtship, although its roles in these neurons during development or adulthood were not yet discriminated.
Taken together, the above results demonstrate distinct roles of fruM expression during a critical developmental period for the manifestation of courtship behaviors and adulthood for inhibiting male–male courtship (Figure 4A), and further reveal that different fruM expression levels and patterns determine courtship modes, indicative of functional flexibility of the fruM-expressing sex circuitry tuned by fruM function (Figure 4B).
Figure 4
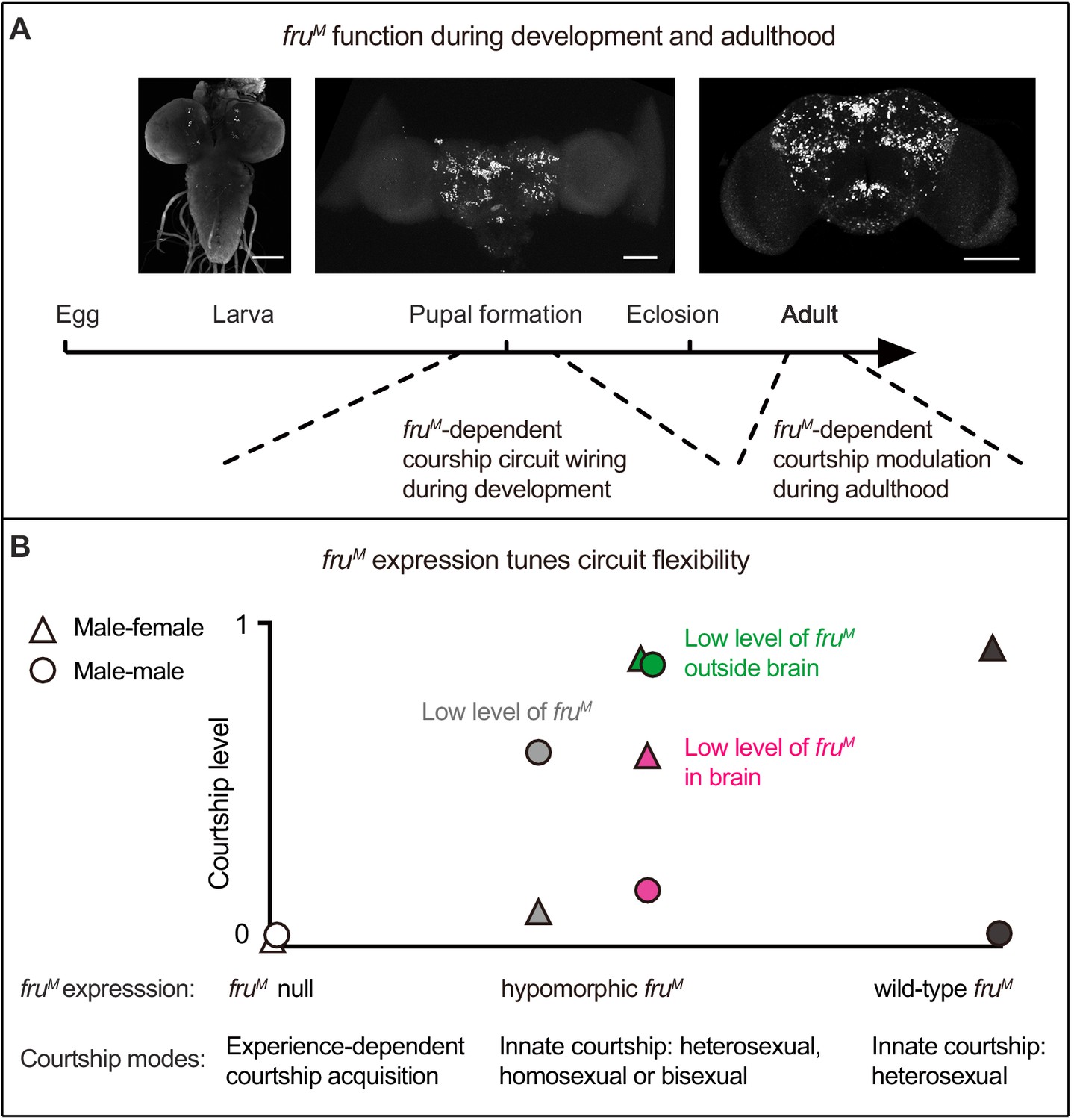
A summary of fruM function in male courtship.
(A) FruM is required during pupation for neuronal development and possibly circuit wiring that builds the potential for innately female-directed courtship, while its function during adulthood is involved in inhibiting male–male courtship. Anti-V5 signals indicate FruM expression in larva, pupa and adult males (from left to right). Scale bars, 100 μm. (B) The sex circuitry without fruM or with different levels/patterns of fruM has different properties such that males would have experience-dependent courtship acquisition, or innate courtship but with different sexual orientation (heterosexual, homosexual, or bisexual). Such flexibility of the sex circuitry is tuned by different fruM expression. Triangles and circles represent corresponding fruM levels and courtship levels (triangles: male–female courtship; circles: male–male courtship). Gray indicates systemic low level of fruM; green and magenta indicate spatially low level of fruM.
Discussion
Previous findings show that fruM expression commences at the wandering third-instar larval stage, peaks at the pupal stage, and thereafter declines but does not disappear after eclosion (Lee et al., 2000), which suggests that fruM may function mainly during development for adult courtship behavior despite of no direct evidence. Here we temporally knocked down fruM expression in different developmental stages for 2 days and found that males with fruM knocked down during pupation rarely courted, while males with fruM knocked down during adulthood courted normally toward females. This is the first direct evidence that fruM is required during development but not adulthood for female-directed courtship behavior. A caveat of these experiments is that while fruM expression is effectively knocked down upon 2 day induction of fruM microRNA, it is not restored acutely after transferring to permissive temperature, although it is restored in adulthood if induction of fruM microRNA was performed at earlier stages (stages 1–5). Such a caveat does not compromise the above conclusion as knocking down fruM during pupation (stage 5) almost eliminated male courtship while knocking down at later stages have minor or no effect on male courtship. Consistent with these behavioral findings, knocking down fruM during stages 5 and 6, but not later stages, results in developmental defect in the gustatory receptor neurons innervating VNC.
In addition to the role of fruM during development to specify female-directed courtship, we also found a role of fruM during adulthood in suppressing male–male courtship, as males with fruM knocked down or tra overexpressed during adulthood displayed enhanced male–male courtship or male chaining behaviors. Note that a previous study found that removal of transformer 2 (tra2) specifically during adulthood using a temperature sensitive tra2 allele induced 8 of 96 females to show male-type courtship behaviors (Belote and Baker, 1987), which suggests that expression of FRUM and DSXM (by removal of tra2 function in females) during adulthood is sufficient to masculinize CNS to some extent and induce a small fraction of females to display male-like courtship behaviors. Recent studies also found that fruM expression in the Or47b-expressing olfactory receptor neurons as well as their neuronal sensitivity depend on social experiences during adulthood (Hueston et al., 2016; Sethi et al., 2019). Based on all these findings, we propose that fruM expression during pupation is crucial for neuronal development and reconstruction of adult sex circuitry that allows innate courtship toward females, and its expression during adulthood may be activity dependent in at least some neurons and modulates some aspects of courtship (e.g., inhibits male–male courtship). Thus, there are at least two separate mechanisms that fruM contributes to the sex circuitry, one during a critical developmental period to build the female-directed innate courtship into that circuitry, and the other during adulthood to modulate neuronal physiology in an experience-dependent manner.
Most importantly, we revealed striking flexibility of the fly sex circuitry by manipulating fruM expression. We listed four cases with fruM manipulation here for comparison: (1) males with a sex circuitry having wild-type fruM function have innate heterosexual courtship, as they court readily toward females, but do not court males no matter how long they meet; (2) males with a sex circuitry having no fruM function lose the innate courtship ability, but have the potential to acquire courtship toward males, females, and even other species in an experience-dependent manner; (3) males with a sex circuitry having limited fruM expression (e.g., 40%) have innate homosexual courtship, as they court readily toward other males, but rarely court females; (4) males with a sex circuitry having limited fruM expression outside brain (but intact fruM expression in brain) are innately bisexual, as they court equally toward females or males. Although previous studies found that different fruM alleles (e.g., deletions, inversions, or insertions related to fru) showed very different courtship abnormalities (Anand et al., 2001; Villella et al., 1997), it was very hard to link fruM function to the flexibility of sex circuitry and often seen as allele-specific or background-dependent phenotypes. Our study using relatively simple genetic manipulations that generate dramatical different courtship behaviors promoted us to speculate a different view about the role of fruM: instead of simply being a master gene that controls all aspects of male courtship, fruM is not absolutely necessary for courtship, but changes the wiring of the sex circuitry during development such that the sex circuitry may function in very different ways, ranging from innately heterosexual, homosexual, bisexual, to largely experience-dependent acquisition of the behavior. Such flexibility of the sex circuitry is tuned by different fruM expression, such that changes of fruM regulatory regions during evolution would easily select a suitable functional mode of the sex circuitry.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | Mouse monoclonal anti-Bruchpilot antibody (nc82) | Developmental Studies Hybridoma Bank | Cat# nc82, RRID:AB_2314866 | IHC (1:50) |
| Antibody | Rabbit polyclonal anti-GFP | Thermo Fisher Scientific | Cat# A-11122, RRID:AB_221569 | IHC (1:1000) |
| Antibody | Donkey polyclonal anti-Rabbit, Alexa Fluor 488 | Thermo Fisher Scientific | Cat# A-21206, RRID:AB_2535792 | IHC (1:500) |
| Antibody | Donkey polyclonal anti-Mouse, Alexa Fluor 555 | Thermo Fisher Scientific | Cat# A-31570, RRID:AB_2536180 | IHC (1:500) |
| Antibody | Mouse monoclonal anti-V5-Tag:DyLight550 | Bio-Rad | Cat# MCA1360D550GA, RRID:AB_2687576 | IHC (1:500) |
| Antibody | Rabbit polyclonal anti-FruM | This study | N/A | IHC (1:200) |
| Plasmid | pCFD4 | Addgene | # 49411 | |
| Plasmid | pHD-DsRed | Addgene | # 51434 | |
| Plasmid | pET-28a | Sigma–Aldrich | # 69864 | |
| Chemical compound, drug | Normal Goat Serum (NGS) | Jackson ImmunoResearch Laboratories | Code# 005-000-121 RRID:AB_2336990 | |
| Chemical compound, drug | Paraformaldehyde (PFA) | Sigma–Aldrich | CAS# 30525-89-4 | 4% PFA in 1× PBS |
| Genetic reagent (D. melanogaster) | fruV5 | This study | N/A | Described below |
| Genetic reagent (D. melanogaster) | UAS-mCD8GFP; fruGAL4 | Stockinger et al., 2005 | N/A | |
| Genetic reagent (D. melanogaster) | UAS-fruMi | Meissner et al., 2016 | N/A | |
| Genetic reagent (D. melanogaster) | UAS-fruMiScr | Meissner et al., 2016 | N/A | |
| Genetic reagent (D. melanogaster) | fruLexA | Mellert et al., 2010 | N/A | |
| Genetic reagent (D. melanogaster) | fru4-40 | Pan and Baker, 2014 | N/A | |
| Genetic reagent (D. melanogaster) | fruSat15 | Pan and Baker, 2014 | N/A | |
| Genetic reagent (D. melanogaster) | fruAJ96u | Pan and Baker, 2014 | N/A | |
| Genetic reagent (D. melanogaster) | ppk23-GAL4 | Thistle et al., 2012 | N/A | |
| Genetic reagent (D. melanogaster) | Otd-Flp | Asahina et al., 2014 | N/A | |
| Genetic reagent (D. melanogaster) | tub-GAL80ts | Bloomington Drosophila Stock Center | BDSC_7019 | |
| Genetic reagent (D. melanogaster) | tub>GAL80> | Bloomington Drosophila Stock Center | BDSC_38881 | |
| Genetic reagent (D. melanogaster) | tub>stop>GAL80 | Bloomington Drosophila Stock Center | BDSC_39213 | |
| Genetic reagent (D. melanogaster) | UAS-traF | Bloomington Drosophila Stock Center | BDSC_4590 | |
| Genetic reagent (D. melanogaster) | R15A01-AD | Bloomington Drosophila Stock Center | BDSC_68837 | |
| Genetic reagent (D. melanogaster) | R71G01-DBD | Bloomington Drosophila Stock Center | BDSC_69507 | |
| Software, algorithm | ImageJ | National Institutes of Health | https://imagej.nih.gov/ij/ | |
| Software, algorithm | Prism 8 | GraphPad | https://www.graphpad.com/ |
Fly stocks
Request a detailed protocolFlies were maintained at 22 or 25°C in a 12 hr:12 hr light:dark cycle. Canton-S flies were used as the wild-type strain. Other stocks used in this study include the following: fruGAL4 (Stockinger et al., 2005), fruV5 (this study), UAS-fruMi (attp40), UAS-fruMi (attp2), and UAS-fruMiScr (attp2) (Meissner et al., 2016), fruLexA, fru4-40, fruAJ96u3, and fruSat15 (Pan and Baker, 2014), ppk23-GAL4 (Thistle et al., 2012), P1-splitGAL4 (R15A01-AD; R71G01-DBD) (Zhang et al., 2018), and Otd-Flp (Asahina et al., 2014). UAS-traF (BL#4590), tub-GAL80ts (BL#7019), tub>GAL80> (BL#38881), and tub>stop>GAL80 (BL#39213) were from Bloomington Drosophila Stock Center.
Generation of fruV5
Request a detailed protocolfruV5 was generated by fusing V5 tag in frame with the start codon of fruP1. To generate the fruV5 knock-in line, two gRNAs (gRNA1: 5′-GCCATTAGTGTCGCGGTGCG-3′; gRNA2: 5′-GCGGCCGCGCGAGTCGCCGC-3′) against fru were inserted into pCFD4 vector (Addgene #49411) to induce DNA break near the start codon of fruP1. Then, ~2.1 kb 5′ homologous arm was incorporated into the 5′ MCS of pHD-DsRed (Addgene #51434) through Gibson assembly (digested with NheI and NdeI). To insert V5 tag after the start codon of fruP1, ~2.4 kb 3′ homologous arm was divided into two fragments and amplified separately. These two fragments including the V5 sequence were then subcloned into the 3′ MCS of pHD-DsRed (containing the above 5′ homologous arm) through Gibson assembly (digested with BglII and XhoI). The modified pCFD4 and pHD-DsRed plasmids were injected into vas-cas9 embryos. Successful knock in was selected by 3xP3-DsRed (DsRed-positive eyes) and confirmed by PCR followed by sequencing. The verified knock-in line was balanced and crossed to hs-Cre flies to remove the 3xP3-DsRed marker.
Generation of anti-FruM antibody
Request a detailed protocolThe rabbit polyclonal antibody against FruM was generated by ABclonal (Wuhan, China). In brief, the fragment of fru gene encodes the N-terminal 101 amino acids, starting with MMATSQDYFG and ending in SPRYNTDQGA, was cloned into expression vector pET-28a (Sigma–Aldrich, #69864). The 101 amino acids are only present in male-specific Fru proteins (FruM) from fruP1. A SUMO-tagged FruM fusion antigen was synthesized from bacteria, purified, and used to immunize a rabbit. The anti-FruM antibody was affinity purified.
Courtship and chaining assays
Request a detailed protocolFor the single-pair courtship assay, the tester males and target flies (4–8 days old) were gently aspirated into round two-layer chambers (diameter: 1 cm; height: 3 mm per layer) and were separated by a plastic transparent barrier that was removed ~30 min later to allow courtship test. Courtship index (CI), which is the percentage of observation time a fly performs any courtship step, was used to measure courtship to female targets or between two males. Paired male–male courtship used two males of the same genotype but focused on the male fly that first initiated courtship (courtship of the initiator to the other). All tester flies were single housed if not otherwise mentioned. Each test was performed for 10 min.
For male chaining assay, tester males (4–8 days old) were loaded into large round chambers (diameter: 4 cm; height: 3 mm) by cold anesthesia. Tests were performed daily for four consecutive days (3 hr after grouping as day 0, then days 1–3). For chaining behavior in Figure 2C, flies were only tested after grouping together for 3 days. Chaining index (ChI), which is the percentage of observation time at least three flies engaged in courtship together, was used to measure courtship in groups of eight males.
To generate males with fruM knocked down only for 2 days during development or adulthood, we raised tub-GAL80 ts/+; fruGAL4/UAS-fruMi flies at 18°C and transferred these flies to fresh food vials every 2 days. In this way, we generated tub-GAL80 ts/+; fruGAL4/UAS-fruMi flies at nine different stages from embryos (stage 1) to newly eclosed adults (stage 9), with wandering larvae at stage 5 and early pupas at stage 6. We then transferred all these flies to a 30°C incubator allowing fruM knockdown for 2 days, then placed all flies back to 18°C until courtship test at adult.
Quantitative real-time PCR
Request a detailed protocolTotal RNA was extracted from ~15 male flies with TRIzol (15596026, Invitrogen), according to the manufacturer’s instructions. The cDNA was synthesized using Prime Script reagent kit (18091050, Invitrogen). Quantitative PCR was performed on LightCycler 96 Real-Time PCR System (Roche) using AceQ qPCR SYBR Green Master Mix (Q121-02, Vazyme). Actin was used as control for normalization. The primers used were as follows: Actin (forward: 5′- CAGGCGGTGCTTTCTCTCTA-3′; reverse: 5′-AGCTGTAACCGCGCTCAGTA-3′), fru P1 promotor (forward: 5′-GTGTGCGTACGTTTGAGTGT-3′; reverse: 5′-TAATCCTGTGACGTCGCCAT-3′), and fru P4 promotor (forward: 5′-TGTATAGCGGCAACTGAACC-3′; reverse: 5′-CCGGTCAAATTTGTGGGATG-3′).
Immunohistochemistry
Request a detailed protocolWe dissected brains and ventral nerve cords of males in defined developmental stages (e.g., Figure 1E–H) or 5–7 days old males in Schneider’s insect medium (Thermo Fisher Scientific, Waltham, MA) and fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 30 min at room temperature. After washing four times in 0.5% Triton X-100% and 0.5% bovine serum albumin [BSA] in PBS (PAT), tissues were blocked in 3% normal goat serum (NGS) for 60 min, then incubated in primary antibodies diluted in 3% NGS for ~24 hr at 4°C, washed (4× 15 min) in PAT at room temperature, and incubated in secondary antibodies diluted in 3% NGS for ~24 hr at 4°C. Tissues were then washed (4× 15 min) in PAT and mounted in Vectorshield (Vector Laboratories, Burlingame, CA) for imaging. Primary antibodies used were rabbit anti-FruM (1:200; this study), mouse anti-V5-Tag:DyLight550 (1:500; MCA1360D550GA, Bio-Rad, Hercules, CA), rabbit anti-GFP (1:1000; A11122, Invitrogen, Waltham, MA), and mouse anti-Bruchpilot (1:50; nc82, Developmental Studies Hybridoma Bank, Iowa City, IA). Secondary antibodies used were donkey anti-mouse IgG conjugated to Alexa 555 (1:500, A31570, Invitrogen) and donkey anti-rabbit IgG conjugated to Alexa 488 (1:500, A21206, Invitrogen). Samples were imaged at 10× or 20× magnification on a Zeiss 700 confocal microscope and processed with ImageJ.
Statistics
Experimental flies and genetic controls were tested at the same condition, and data are collected from at least two independent experiments. Statistical analysis is performed using GraphPad Prism and indicated inside each figure legend. Data presented in this study were first verified for normal distribution by D’Agostino–Pearson normality test. If normally distributed, Student’s t test is used for pairwise comparisons, and one-way ANOVA is used for comparisons among multiple groups, followed by Tukey’s multiple comparisons. If not normally distributed, Mann–Whitney U test is used for pairwise comparisons, and Kruskal–Wallis test is used for comparisons among multiple groups, followed by Dunn’s multiple comparisons.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files. Source data files have been provided for Figures 1, 2, 3, Figure 3-figure supplement 1, 2 and 4.
References
-
Molecular genetic dissection of the sex-specific and vital functions of the Drosophila melanogaster sex determination gene fruitlessGenetics 158:1569–1595.
-
Sex and the single cell I. On the action of major loci affecting sex determination in Drosophila melanogasterGenetics 94:383–423.
-
Sexual dimorphism in the fly brainCurrent Biology 20:1589–1601.https://doi.org/10.1016/j.cub.2010.07.045
-
Sex mysteries of the fly courtship master regulator fruitlessFrontiers in Behavioral Neuroscience 13:245.https://doi.org/10.3389/fnbeh.2019.00245
-
The mode of action of fruitless: is it an easy matter to switch the sex?Genes, Brain and Behavior 19:e12606.https://doi.org/10.1111/gbb.12606
-
Extended reproductive roles of the fruitless gene in Drosophila melanogaster revealed by behavioral analysis of new fru mutantsGenetics 147:1107–1130.
-
Genes and circuits of courtship behaviour in Drosophila malesNature Reviews Neuroscience 14:681–692.https://doi.org/10.1038/nrn3567
-
Cellular organization of the neural circuit that drives Drosophila Courtship BehaviorCurrent Biology : CB 20:1602–1614.https://doi.org/10.1016/j.cub.2010.08.025
Article and author information
Author details
Funding
National Natural Science Foundation of China (31970943)
- Yufeng Pan
National Natural Science Foundation of China (31700905)
- Qionglin Peng
National Natural Science Foundation of China (31622028)
- Yufeng Pan
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank the Bloomington Drosophila Stock Center for fly stocks. This work was supported by grants from National Key R and D Program of China (2019YFA0802400), the National Natural Science Foundation of China (31970943, 31622028, and 31700905), and the Jiangsu Innovation and Entrepreneurship Team Program.
Copyright
© 2021, Chen et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,543
- views
-
- 575
- downloads
-
- 29
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 29
- citations for umbrella DOI https://doi.org/10.7554/eLife.59224
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
fruitless tunes functional flexibility of courtship circuitry during development
eLife 10:e59224.
https://doi.org/10.7554/eLife.59224




{kind=link}
{kind=link}
{kind=link}
{kind=link}