Mitochondrial MICOS complex genes, implicated in hypoplastic left heart syndrome, maintain cardiac contractility and actomyosin integrity
- Development, Aging and Regeneration Program, Center for Genetic Disorders & Aging Research, Sanford Burnham Prebys Medical Discovery Institute, United States
- Department of Bioengineering, Sanford Consortium for Regenerative Medicine, UCSD, School of Medicine, United States
- Cardiovascular Genetics Research Laboratory, Mayo Clinic, United States
- Division of Computational Biology, Department of Quantitative Health Sciences, Mayo Clinic, United States
- Department of Pediatrics, UCSD School of Medicine, La Jolla, Rady’s Hospital MC 5004, United States
- Center for Regenerative Medicine, Division of Pediatric Cardiology, Department of Pediatric and Adolescent Medicine, Division of General Internal Medicine, Department of Molecular and Pharmacology and Experimental Therapeutics, Mayo Clinic, United States
- Department of Cardiovascular Medicine, Division of Pediatric Cardiology, Department of Pediatric & Adolescent Medicine, Cardiovascular Genetics Research Laboratory, Mayo Clinic, United States
Abstract
Hypoplastic left heart syndrome (HLHS) is a severe congenital heart disease (CHD) with a likely oligogenic etiology, but our understanding of the genetic complexities and pathogenic mechanisms leading to HLHS is limited. We performed whole genome sequencing (WGS) on 183 HLHS patient-parent trios to identify candidate genes, which were functionally tested in the Drosophila heart model. Bioinformatic analysis of WGS data from an index family of a HLHS proband born to consanguineous parents prioritized 9 candidate genes with rare, predicted damaging homozygous variants. Of them, cardiac-specific knockdown (KD) of mitochondrial MICOS complex subunit dCHCHD3/6 resulted in drastically compromised heart contractility, diminished levels of sarcomeric actin and myosin, reduced cardiac ATP levels, and mitochondrial fission-fusion defects. These defects were similar to those inflicted by cardiac KD of ATP synthase subunits of the electron transport chain (ETC), consistent with the MICOS complex’s role in maintaining cristae morphology and ETC assembly. Five additional HLHS probands harbored rare, predicted damaging variants in CHCHD3 or CHCHD6. Hypothesizing an oligogenic basis for HLHS, we tested 60 additional prioritized candidate genes from these patients for genetic interactions with CHCHD3/6 in sensitized fly hearts. Moderate KD of CHCHD3/6 in combination with Cdk12 (activator of RNA polymerase II), RNF149 (goliath, E3 ubiquitin ligase), or SPTBN1 (β-Spectrin, scaffolding protein) caused synergistic heart defects, suggesting the likely involvement of diverse pathways in HLHS. Further elucidation of novel candidate genes and genetic interactions of potentially disease-contributing pathways is expected to lead to a better understanding of HLHS and other CHDs.
Editor's evaluation
In the revised version, Birker et al. add experiments in flies and human iPSC-derived cardiomyocytes and add new text that further supports their central claim that mutations in MICOS complex components mediate HLHS by reducing sarcomere integrity and heart contractility. The ability to go from identifying variants by whole genome sequencing in HLHS patients to generating oligogenic animal models to test whether these variants produce cardiac phenotypes is well demonstrated here and highlights the importance of model organisms in disease research. Overall, the manuscript is improved, and the data support the claims.
https://doi.org/10.7554/eLife.83385.sa0Introduction
Hypoplastic left heart syndrome (HLHS) is a birth defect that accounts for 2–4% of congenital heart defects (CHDs), equal to 1000–2000 HLHS births in the United States per year. HLHS has been proposed to be caused by genetic, epigenetic, or environmental factors (Crucean et al., 2017; Liu et al., 2017; Yagi et al., 2018; Grossfeld et al., 2019). The severe cardiac characteristics of HLHS include aortic and mitral stenosis or atresia, and reduced size of the left ventricle and aorta; however, there is a spectrum of cardiac phenotypes that can underly HLHS pathophysiology (Theis et al., 2015a; Crucean et al., 2017; Mussa and Barron, 2017; Grossfeld et al., 2019). If not treated with reconstructive heart surgeries or cardiac transplantation, infants born with HLHS will not survive (Grossfeld et al., 2019). To date, the standard treatment for this disease is a three-stage surgical procedure, which begins neonatally and aims overall to achieve right ventricle-dependent systemic circulation and deliver oxygen-poor blood more directly to the lungs (Mussa and Barron, 2017). Although the surgical procedures correctly divert left ventricular function to the right ventricle, there is a subgroup of HLHS patients who are at risk of latent heart failure, which is often preceded by reduced ejection fraction (Altmann et al., 2000; McBride et al., 2008; Theis et al., 2015b).
Although several studies have examined the molecular underpinnings of HLHS, the number of genes associated with this disease is small (e.g. NKX2-5, NOTCH1, ETS1, MYH6, LRP2, and CELSR1), and they are not yet conclusively determined as causal for HLHS (Garg et al., 2005; Ye et al., 2010; Kobayashi et al., 2014; Theis et al., 2015a; Tomita-Mitchell et al., 2016; Theis et al., 2020; Theis, 2021; Theis et al., 2022). Defining pathogenic mechanisms has proved elusive given the oligogenic complexity of HLHS. Overall, there is a great need to functionally evaluate newly emerging HLHS candidate genes to understand how they may contribute to the molecular, cellular, and morphological processes underlying HLHS.
Drosophila is well-suited for modeling genetic underpinnings of CHDs: many of the genes and gene programs found in the Drosophila heart are evolutionarily conserved, including a core set of cardiogenic transcription factors and inductive factors (e.g. Nkx2-5/tinman) (Bodmer, 1995; Cripps and Olson, 2002; Bier and Bodmer, 2004; Bodmer and Frasch, 2010; Ahmad, 2017), approximately 75% of known human disease-causing genes having fly orthologs (Bodmer and Frasch, 2010; Pandey and Nichols, 2011; Ugur et al., 2016), and the developing mammalian and Drosophila hearts share developmental similarities, such as their origin within the mesoderm.
Mitochondria have been postulated to play a critical role in HLHS pathogenesis. For example, a recent study reported that cardiomyocytes derived from iPSCs of HLHS patients (iPSC-CM), who later developed right ventricular failure, had reduced mitochondrial concentration, ATP production, and contractile force (Paige et al., 2020). This study revealed downregulated expression of genes involved in mitochondrial processes, such as ATP synthesis coupled electron transport. Another study of HLHS patient-derived iPSC-CMs revealed reduced mitochondrial size, number, and malformed mitochondrial inner membranes using transmission electron microscopy (Yang et al., 2017). Similarly, an HLHS mouse model with Sap130 and Pcdha9 mutations showed mitochondrial defects manifested as reduced cristae density and smaller mitochondrial size (Liu et al., 2017). Despite a lack of understanding of the exact mitochondrial mechanisms underlying HLHS pathogenesis, recent experimental and bioinformatic data suggest an underlying role of mitochondria in HLHS.
Here, a cohort of 183 HLHS proband-parent trios underwent whole genome sequencing (WGS) to identify candidate genes, including a prioritized consanguineous family where genes harboring rare, predicted damaging homozygous variants were investigated (Theis and Olson, 2022). Among the resulting candidate HLHS genes tested in Drosophila, cardiac-specific knockdown (KD) of Chchd3/6 (coiled-coil-helix-coiled-coil-helix-domain-containing protein 6) of the MICOS (mitochondrial contact site and cristae organization system) complex exhibited severe heart structure and function defects. The MICOS complex is an eight-subunit complex in mammals (five in Drosophila) located in the inner mitochondrial membrane that is necessary to maintain cristae morphology and ATP production. It is closely associated and interacts with SAMM50 (sorting and assembly machinery, CG7639), which is located in the outer mitochondrial membrane (Ott et al., 2012; Kozjak-Pavlovic, 2017). The MICOS complex’s role in cardiac development and functional homeostasis is not known but is likely important for efficient ATP production. We observed reduced contractility upon cardiac-specific Chchd3/6 KD, diminished sarcomeric Actin and Myosin levels, as well as severe mitochondrial morphology defects, which manifested as fragmented and aggregated structures. Similar phenotypes were observed upon cardiac KD of other MICOS complex genes, as well as other mitochondrial genes such as ATP synthase (complex V), specifically ATP synthase B and β. We also found significantly diminished proliferation of human induced pluripotent stem cell (iPSC)-derived ventricular-like cardiomyocytes (VCMs) upon KD of MICOS genes. Finally, a family-based candidate gene interaction screen in Drosophila revealed three genes that genetically interact with Chchd3/6: Cdk12 (activator RNA polymerase II activator), RNF149 (goliath, gol, E3 ubiquitin ligase), SPTBN1 (β Spectrin, β-Spec, scaffolding protein). In summary, Chchd3/6 and other components important for mitochondrial homeostasis were identified as critical for establishing and maintaining cardiac structure and function, and likely contribute to HLHS and/or latent heart failure following surgical palliation.
Results
Family phenotype
Family 11 H is of white ancestry and comprised of a male with HLHS, his parents, and two siblings, they were all phenotypically characterized by echocardiography and underwent WGS. A homozygous recessive disease mode of inheritance was postulated due to reported consanguinity between the mother and father and absence of structural and myopathic heart disease in the parents (Figure 1A). The siblings also had normal echocardiograms. The 11 H proband had latent decline of right ventricular ejection fraction several years after surgical palliation. In addition to HLHS, he was diagnosed with developmental delay, cerebral and cerebellar atrophy, white matter loss, decreased muscle mass, and a body mass index <1%, traits that have previously been related to mitochondrial dysfunction (Alston et al., 2017; Romanello and Sandri, 2016).
Figure 1 with 1 supplement see all
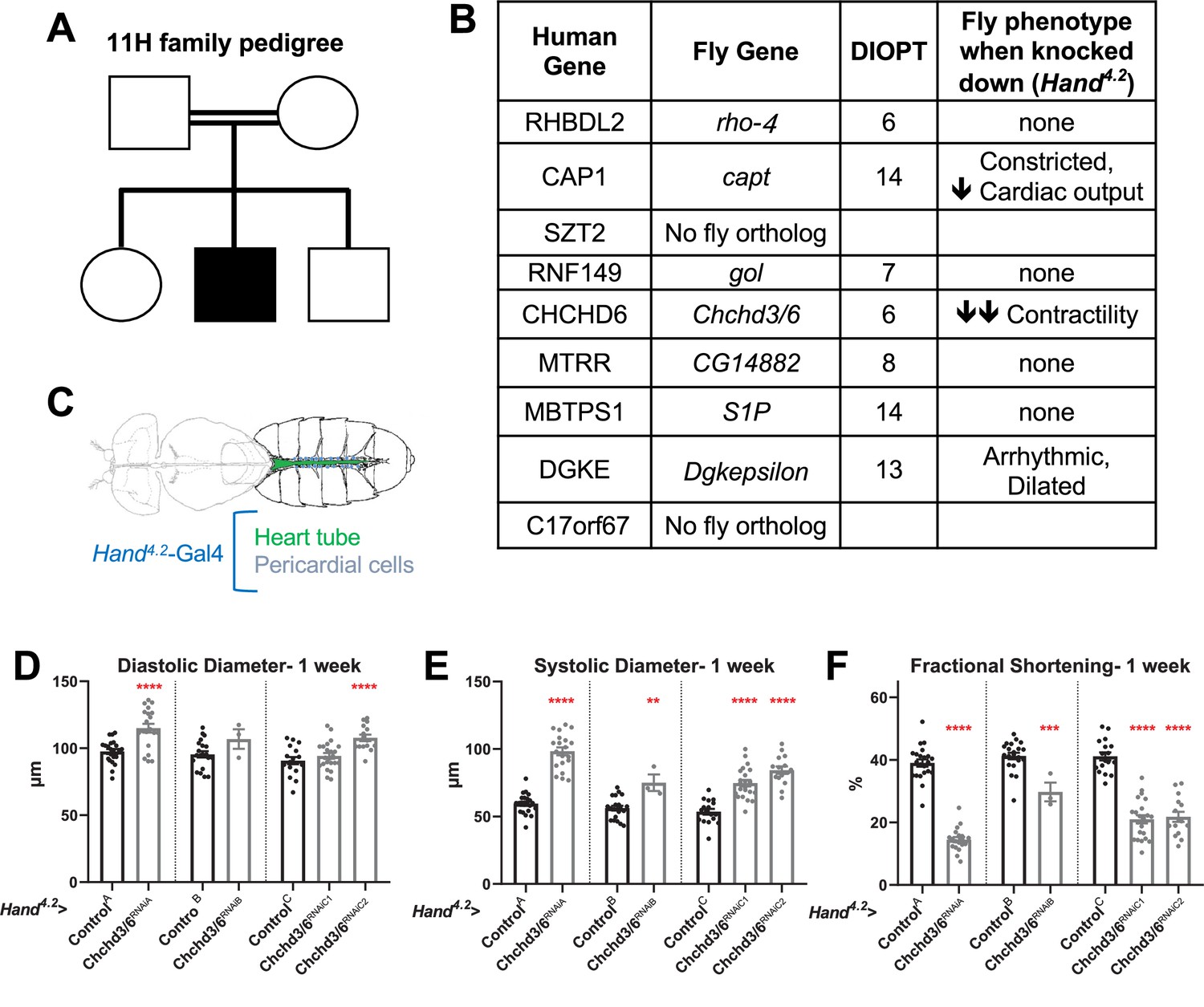
Prioritization of CHCHD6 in HLHS proband and its Drosophila ortholog Chchd3/6.
(A) Pedigree of index family 11 H. The family includes consanguineous parents (denoted by double horizontal lines) without cardiac defects, one son with HLHS (proband), and two siblings without cardiac defects. (B) List of 9 candidate genes derived from proband 11 H with corresponding Drosophila orthologs. Orthology based on DIOPT score. Conserved Drosophila candidate HLHS genes were knocked down individually in the Drosophila heart using the Hand4.2-Gal4 driver. The functional phenotypes listed were significantly different relative to ControlA or ControlB and were measured in 1-week-old female Drosophila hearts. (C) Schematic of Drosophila highlighting the abdominal region which includes the heart tube and flanking pericardial cells, where the Hand4.2-Gal4 driver is expressed. Image adapted from Figure 1A of Xie et al., 2013. (D) End-Diastolic diameter (EDD), (E) End-systolic diameter (ESD), and (F) fractional shortening (FS) from 1-week-old female Hand4.2-Gal4>Chchd3/6 flies.
-
Figure 1—source data 1
List of 9 candidate genes derived from proband 11 H with corresponding Drosophila orthologs.
Orthology based on DIOPT score. Conserved Drosophila candidate HLHS genes were knocked down individually in the Drosophila heart using the Hand4.2-Gal4 driver. The functional phenotypes listed were significantly different relative to ControlA or ControlB and were measured in 1-week-old female Drosophila hearts.
- https://cdn.elifesciences.org/articles/83385/elife-83385-fig1-data1-v2.xlsx
Whole genome sequencing and bioinformatics analysis of 11H family
Array comparative genomic hybridization ruled out a chromosomal deletion or duplication in the proband. WGS was carried out on genomic DNA samples from the five family members, based on paired-end reads that passed quality control standards; 99.4% of the reads mapped to the genome. After marking and filtering out duplicate reads, over 91% of the hg38 human reference genome had coverage. The average depth across the genome was 63 X and an average of 89% of the genome demonstrated a minimal read depth of 20 reads. Filtering for rare variants that were homozygous in the HLHS proband revealed nine candidate genes. Three genes had a missense variant (SZT2, MTRR, MBTPS1) whereas the remaining six genes were found to have a non-coding variant within the promoter (CAP1, DGKE), 5’ untranslated region (RHBDL2, RNF149, C17orf67), or intron (CHCHD6)(Marian et al., 2011). While six of the variants were also found to be homozygous in an unaffected sibling, the associated candidate genes were not excluded from downstream analyses based on the postulated oligogenic nature of HLHS, and incomplete penetrance of individual variants, as observed in a digenic mouse model (Yagi et al., 2018).
Candidate HLHS gene knockdown in Drosophila reveals requirement for Chchd3/6 in establishing cardiac structure and function
To test whether the HLHS candidates had significant requirements in the heart, we utilized the established Drosophila heart model and cardiac-specific RNAi KD. First, the nine candidate genes were assigned their respective Drosophila homologs; seven out of nine of the human HLHS candidate genes had Drosophila orthologs (Figure 1B; Hu et al., 2011). The Drosophila Gal4-UAS system (Brand and Perrimon, 1993) was used to test candidate genes for their role in heart function using temporal and/or spatial KD via RNAi. The Hand4.2-Gal4 driver was used for initial screening because it is a strong post-mitotic and heart-specific driver, which is expressed throughout life in the cardiomyocytes (CMs) and pericardial cells (PCs) (Han and Olson, 2005; Han et al., 2006; Figure 1C). Three-week-old (mid-adult stage) female flies were used to test the seven candidate genes. Hand4.2-Gal4 KD of capt (actin binding protein, negatively regulating actin filament assembly), Dgkepsilon (Diacyl glycerol kinase, DGKE), and Chchd3/6 (Mitochondrial inner membrane protein of the MICOS complex, required for fusion) produced defects in the fly hearts, such as reduced cardiac output, reduced fractional shortening, and arrhythmicity (Figure 1B). Of those, Chchd3/6 KD gave the most severe cardiac defects with strongly reduced fractional shortening, a measure of cardiac contractility. Systolic rather than diastolic diameter was increased, which suggests systolic dysfunction (Figure 1—figure supplement 1A–C). Since reduced contractility was previously shown in animals with reduced mitochondrial gene expression, we hypothesized Chchd3/6 KD may reduce contractility via a role in mitochondrial function (Bhandari et al., 2015; Martínez-Morentin et al., 2015; Tocchi, 2015).
To test how early the cardiac phenotype of Chchd3/6 KD manifests in adult stages, 1-week-old Hand4.2-Gal4 KD of Chchd3/6 flies were examined, using several independent RNAi lines for Chchd3/6. These flies also had reduced fractional shortening, that is, reduced contractility due to systolic dysfunction (Figure 1D–F). This phenotype was observed in cardiac assays of intact flies (see Materials and methods; Figure 1—figure supplement 1D–F), as well as in the semi-intact adult heart preparation that lacks neuronal inputs (SOHA; Fink et al., 2009). To further validate a cardiac-specific role for Chchd3/6, as opposed to non-autonomous effects from other tissues, we performed KD of Chchd3/6 using Dot-Gal4 (expressed in pericardial cells, PC, which also express Hand), Mef2 (Myocyte enhancer factor 2)-Gal4 (a pan-muscle driver), or elav-Gal4 (a pan-neuronal driver; see Materials and methods). A large reduction in fractional shortening was only observed with the pan-muscle driver that includes cardiac muscle, but not with the PC or neuronal drivers, confirming a cardiomyocyte-autonomous effect (Figure 1—figure supplement 1J–L). Both Chchd3/6RNAiA and Chchd3/6RNAiB lines had the same predicted off-target gene, Duox but Hand4.2-Gal4 driven KD of Duox had no effect on fractional shortening, confirming that the cardiac effects were due to Chchd3/6 KD (Figure 1—figure supplement 1M).
Temporal requirements for Chchd3/6 in maintaining heart function
We next sought to understand if Chchd3/6 has different temporal requirements for its effects on heart structure and function, since hearts of operated HLHS patients often develop reduced ejection fraction and heart failure, including the 11 H proband. First, to assess whether Chchd3/6 could have a role in early heart development, we mined embryonic heart-specific single-cell transcriptomic data (Vogler et al., 2021b) and found that Chchd3/6 was expressed in Drosophila cardioblasts (CBs), along with other cardiogenic factors (tinman, H15, and Hand) (Figure 2A). Next, we analyzed Chchd3/6 mutant embryos for cardiac phenotypes. Late-stage 16–17 ChchdD1 /ChchdDefA trans-heterozygous embryos were stained for Mef2 (early mesoderm/muscle-specific transcription factor) and Slit (secreted protein in the heart lumen) but did not exhibit overt cardiac specification defects (Figure 2B). We used the tinD-Gal4 driver (tinman enhancer D Yin et al., 1997; Xu et al., 1998) to test whether Chchd3/6 KD in the dorsal mesoderm (including cardiac mesoderm) during embryonic stages 10–12 affects establishment of adult heart function. We reared tinD-Gal4 >Chchd3/6RNAiA flies at 29 °C throughout life to achieve high KD efficiency but did not observe reduced fractional shortening or any other functional defects, relative to controls (Figure 2C and D). Thus, KD of Chchd3/6 in the embryonic cardiac mesoderm is not sufficient to impact later heart function.
Figure 2
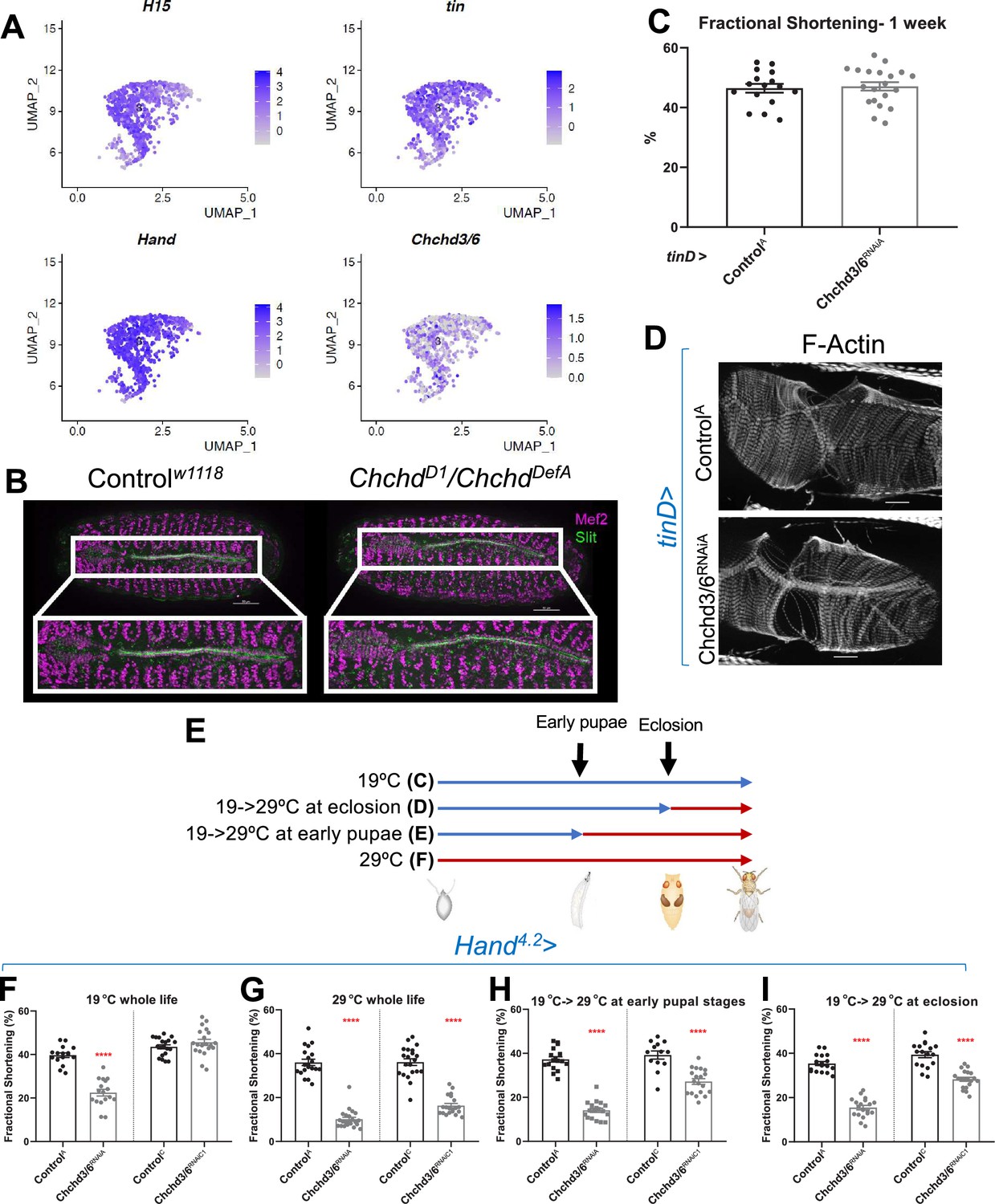
Chchd3/6 expression is important for adult cardiac function around larval stages and early adult stages.
(A) UMAP (uniform manifold approximation and projection) plot from CB-specific single-cell transcriptomics (Vogler, 2021a) showing expression of Chchd3/6 in CBs, as identified by cardiac TFs tin, H15, and Hand. (B) Stage 16–17 embryos (late stage cardiogenesis) were collected from a Chchd3/6 loss of function line (ChchdD1) line crossed to a Chchd3/6 deficiency line (ChchdDefA) and stained for Mef2 (all muscle transcription factor, magenta) and Slit (secreted protein of the lumen, green). 50 µm scale. (C) tinD >ControlA or>Chchd3/6RNAiA were reared at 29 °C and females were filmed and imaged at 1 week of age. (A) tinD >Chchd3/6RNAiA did not have a significant reduction in fractional shortening compared to tinD >ControlA flies. (D) F-actin was unchanged between tinD >ControlA and tinD >Chchd3/6RNAiA flies at 1 week of age; 20 µm scale. (E) Schematic overview of temperature shift experiments. (F–I) Fractional shortening measurements from 1-week-old female flies reared at (F) 19 °C for whole life, (G) at 29 °C for whole life, (H) 19 °C, and moved to 29 °C at early pupal stages, or (I) 19 °C, and moved to 29 °C once eclosed (virgin flies), Unpaired two-tailed t-test, ****p≤0.0001, error bars represent SEM.
To further investigate the temporal requirement of Chchd3/6 during heart development, we made use of the temperature-dependence of Gal4-mediated KDs (less KD efficiency at 19 °C, greater KD efficiency at 29 °C; see Figure 2E for experimental strategy). Hand4.2-Gal4 mediated KD of Chchd3/6RNAiA had strong contractility defects already at 19 °C (Figure 2F). A weaker RNAi KD line Chchd3/6RNAiC1 (see Figure 1D–F) caused no reduction in fractional shortening at 19 °C (Figure 2F), whereas at 29 °C fractional shortening was reduced similarly to the stronger KD line 19 °C. To examine different developmental windows, Hand4.2-Gal4>Chchd3/6RNAiC1 flies were shifted from 19°C to 29°C at either early pupal stages or early adult stages (after eclosion) until 1 week of age when heart function was assessed (Figure 2E). Interestingly, both treatments caused a substantial reduction in fractional shortening, although somewhat less than at 29 °C throughout life (Figure 2G–I). This suggests that Chchd3/6 is not only required during pupal development, but also at adult stages for maintaining robust heart function.
Cardiac knockdown of Drosophila Chchd3/6 results in severe reduction of sarcomeric actin and myosin levels
The strong heart functional defects upon Chchd3/6 KD suggest that the contractile machinery in cardiomyocytes is severely compromised. To probe for contractile abnormalities, we examined several sarcomeric components, including filamentous (F-) Actin, Myosin heavy chain, Obscurin (present at the M-line), α-Actinin (present at the Z-line), and Sallimus (Titin component in flies, localized near the Z-line) (Figure 3A). The intensity of F-actin staining with phalloidin was severely diminished in the working myocardium of Hand4.2-Gal4>Chchd3/6 KD flies (Figure 3B–D, arrows in B). Because Hand4.2-Gal4 expression is less in ostial cardiomyocytes (inflow valves), F-actin staining in ostial sarcomeres was minimally affected, if at all (Figure 3B, arrowheads). Like F-actin staining, Myosin staining was also dramatically diminished (Figure 3E,I) In contrast, Obscurin and α-Actinin staining was only moderately reduced (Figure 3F, G, J and K), and Sallimus staining was unaffected (Figure 3H and L). These findings suggest that loss of Chchd3/6 function did not abrogate the overall sarcomeric organization, but instead differentially affected the abundance of individual sarcomeric proteins. Overall, the strongly diminished F-actin and Myosin levels in cardiac myofibrils is likely responsible for the diminished contractile capacity of the ATP-dependent actomyosin network in Chchd3/6 KD hearts.
Figure 3 with 1 supplement see all
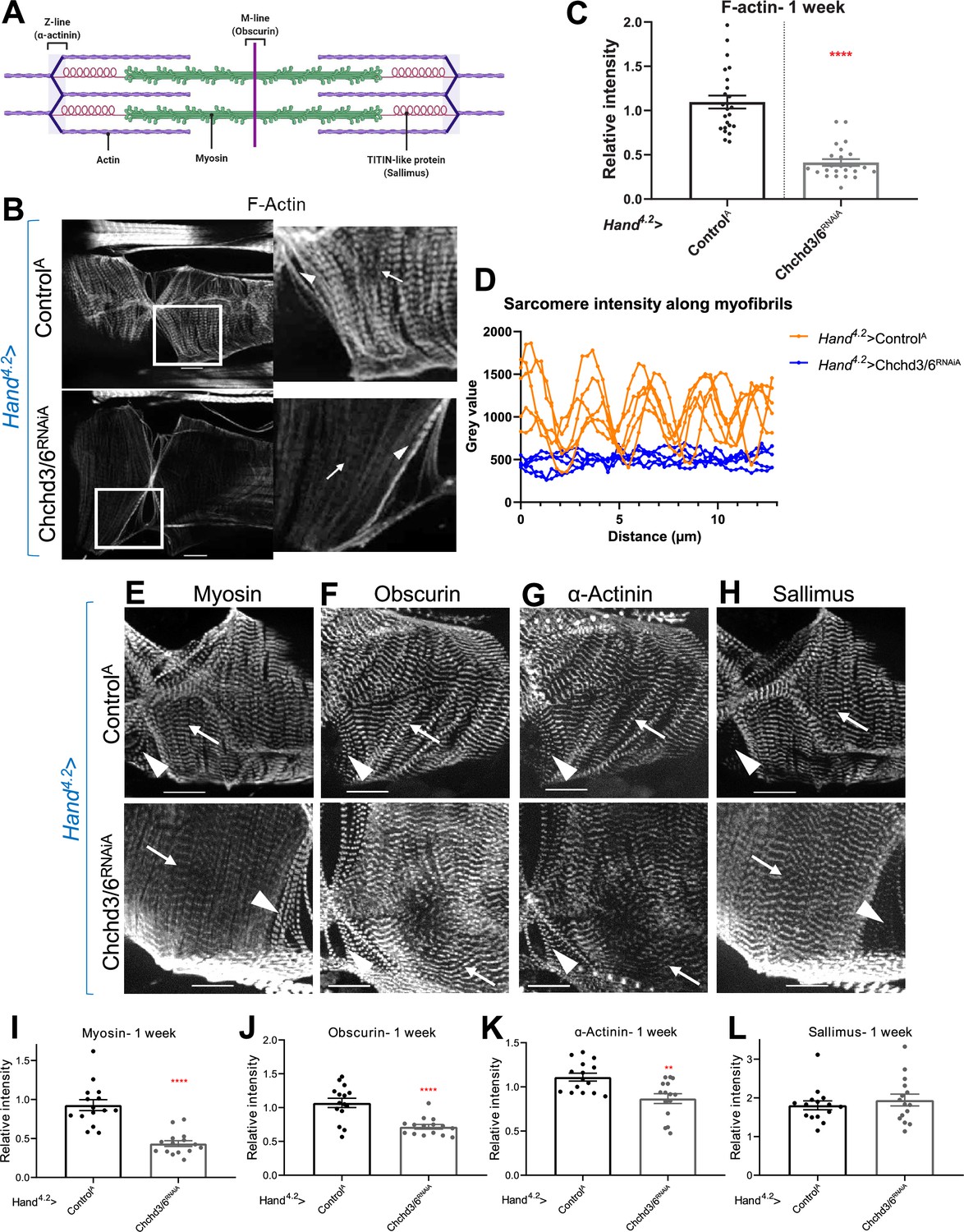
Cardiac tissue from heart-specific Chchd3/6 KD flies exhibit reduced and altered sarcomeric proteins in the myocardial tissue.
(A) Schematic of sarcomeric protein distribution inside myofibrils (image created with BioRender.com). (B) F-actin staining in 1-week-old female Drosophila hearts with Hand4.2-Gal4 KD of Chchd3/6. Arrowheads indicate ostial myofibrils and arrows point to myocardial myofibrils (non-ostial). (C) F-actin intensity measured as mean gray value (gray value/# of pixels) along myocardial myofibrils relative to mean gray value of ostial myofibrils. (D) Mean intensity of F-actin along individual myofibrils. One-week-old Drosophila hearts with Hand4.2-Gal4 driven KD of control or Chchd3/6 stained for antibodies against (E) Myosin, (F) Obscurin, (G) α-Actinin, or (H) Sallimus. Arrowheads indicate ostial myofibrils and arrows point to working cardiomyocyte tissue (non-ostial). (I–L) Mean fluorescence intensity along myocardial myofibrils relative to ostia myofibrils in 1-week-old Hand4.2-Gal4>CHCHD3/6RNAiA adults stained for sarcomeric proteins (I) Myosin, (J) Obscurin, (K) α-Actinin, or (L) Sallimus. Unpaired two-tailed t-test, **p≤0.01, ****p≤0.0001; error bars represent SEM. 20 µm scale.
Actin polymerization components do not mediate sarcomeric actomyosin reduction upon cardiac Chchd3/6 knockdown
Due to the strong reduction of F-actin levels observed with reduced Chchd3/6 expression, we hypothesized that globular (G) to F-actin polymerization was disrupted. If Chchd3/6 KD compromises mitochondrial ATP production in high energy-demanding CMs, the reduced ATP levels could disrupt actin polymerization and lead to reductions in F-actin and other sarcomeric proteins (Carlier et al., 1984; Korn et al., 1987; Carlier, 1998). To test this, we reduced the cardiac expression of several actin polymerizing and depolymerizing genes (Figure 3—figure supplement 1A). Cardiac KD of Arp2/3, gel, Chd64, WASp, and TM1 caused slightly reduced fractional shortening, but not as severe as with Chchd3/6 KD (Figure 3—figure supplement 1A, B). Moreover, cardiac KD of Arp, Chd64, or WASp resulted in substantial myofibrillar disorganization, including gaps, but did not appear to produce the Chchd3/6 KD-like reduction in sarcomeric F-actin levels (WASp example shown in Figure 3—figure supplement 1C). Overall, KD of any of these genes involved in F-actin polymerization could not recapitulate the reduced myocardial F-actin intensity with normal sarcomeric patterning seen with Chchd3/6 KD. Therefore, it appears unlikely that defects in actin polymerization mediate the effects of Chchd3/6 KD.
Chchd3/6 knockdown in flight or heart muscles results in defective mitochondria
Next, we hypothesized that the reduced contractile capacity and altered F-actin and Myosin in Chchd3/6 KD hearts was due to reduced mitochondrial function. We first examined mitochondrial integrity and sarcomeric actin staining in indirect flight muscles (IFMs), since their mitochondria are easily visualized due to their large size. Upon Chchd3/6 KD in IFMs using the pan-muscle driver Mef2-Gal4, we observed reduced F-actin staining and diminished sarcomere pattern definition (Figure 4A), similar to the cardiac phenotype (Figure 3B). This further indicated that the Chchd3/6 KD phenotype is not specific to cardiac tissue, but likely affects all muscles. We then examined mitochondrial integrity upon Chchd3/6 KD in IFMs expressing Mito::GFP (complex IV), and with antibodies against ATP synthase (complex V). Strikingly, Mito::GFP and ATP synthase staining revealed mitochondrial fission-fusion defects (Figure 4B–E), which is suggestive of an imbalance between fusion and fission.
Figure 4 with 2 supplements see all
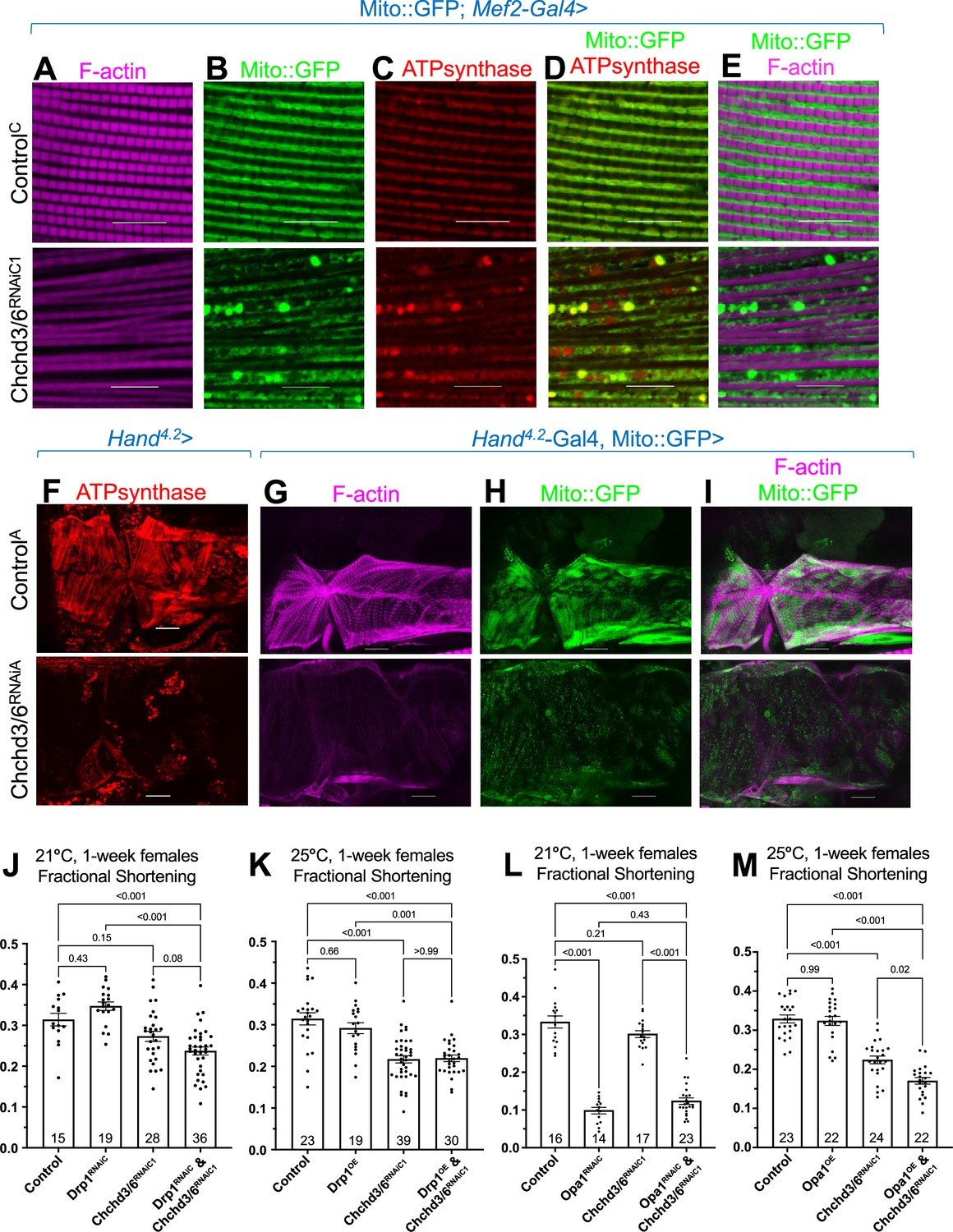
Mitochondrial fission-fusion defects were observed in cardiac Chchd3/6 KD.
(A–E) Visualization of F-actin and mitochondria in Drosophila indirect flight muscles (IFMs). 1–2 day-old male Drosophila IFMs with Mito::GFP; Mef2-Gal4 stained for (A) F-actin, (B) GFP (Mito::GFP (GFP tagged COX8A)), (C) ATP synthase, (D) merged image of B+C, and (E) merged image of A+B,10µm scale. (F) Hand4.2>Chchd3/6RNAiA heart tissue at 1 week of age. (G–I) F-actin and Mito::GFP staining in 1-week-old female hearts using the Hand4.2-Gal4; Mito::GFP driver, 20 µm scale. (J–M) Fractional shortening results are displayed from 1-week-old females with manipulation of mitochondrial fission-fusion genes and Chchd3/6RNAiC1. (J) Chchd3/6 and Drp1 KD at 21 °C had little effect on their own, but in combination caused a reduction in fractional shortening, displaying a significant genetic interaction (Figure 3—figure supplement 1C). (K) At 25 °C Chchd3/6 KD reduced fractional shortening substantially, whereas Drp1 OE by itself or in combination with Chchd3/6RNAiC1 had no effect, thus no genetic interaction was observed (Figure 3—figure supplement 1F). (L) Even at 21 °C, Opa1 KD drastically reduced contractility, which in combination with Chchd3/6 KD slightly improved, which resulted in a significant genetic interaction (Figure 3—figure supplement 1I). (M) At 25 °C, Opa1 OE (M) had no effect, but in combination with Chchd3/6 KD contractility was further reduced significantly, although the interaction p-value did not reach significance (Figure 3—figure supplement 1L). One-way ANOVA with multiple comparisons shows mean with SEM and associated p-values. Sample size is shown at bottom of each bar.
Next, we assayed ATP synthase staining in Hand4.2-Gal4>Chchd3/6RNAiA hearts. We again observed mitochondrial fission-fusion defects, along with reduced F-actin and ATP synthase staining, relative to controls (Figure 4F–I). Furthermore, Hand4.2-Gal4, Mito::GFP >Chchd3/6RNAiA hearts also exhibited mitochondrial fission-fusion defects and reduced intensity of Mito::GFP (Figure 4G–I). Taken together, these data show that Chchd3/6 KD disrupts cardiac mitochondrial morphology.
Mitochondrial fission-fusion genes, Drp1 and Opa1, interacted genetically with Chchd3/6
To explore the role of mitochondrial fission-fusion defects in cardiac Chchd3/6 KD, we conducted genetic interaction experiments. In Drosophila, dynamin related protein 1 (Drp1) promotes mitochondrial fission, and optic atrophy 1 (Opa1) promotes the fusion of the inner mitochondrial membrane (Dorn and Kitsis, 2015). We used KD and overexpression (OE) of these genes in conjunction with a Chchd3/6 sensitizer line we generated, Hand4.2-Gal4,tdtK;Chchd3/6RNAiC1, which at 25 °C, but not at 21 °C, exhibits significant contractility deficits, as measured by fractional shortening (Figure 1). At 21 °C, cardiac KD of Drp1 or Chchd3/6 alone had little effect, except for a slight dilation with Chchd3/6 KD (Figure 4J, Figure 4—figure supplement 1A and B); however, the combined KD reduced contractility considerably, indicative of a genetic interaction (Figure 4J, Figure 4—figure supplement 1C). In addition, F-actin staining in the double KD was diminished and even disorganized, compared to either KD alone (Figure 4—figure supplement 2A). In contrast, Drp1 OE combined with Chchd3/6 KD at 25 °C did not alter the moderate decrease in contractility or F-actin staining (Figure 4K; Figure 4—figure supplement 1F, Figure 4—figure supplement 2B), but did revert the dilation due to Chchd3/6 KD (Figure 4—figure supplement 1D and E).
In contrast to Drp1, Opa1 KD already at 21 °C strongly exhibited systolic dysfunction, which surprisingly was slightly reversed when combined with Chchd3/6 KD, thus indicative of a genetic interaction (Figure 4L, Figure 4—figure supplement 1G–I), possibly due to a dual role of Opa1 (Pernas and Scorrano, 2016). F-actin staining was also diminished by Opa1 KD (Figure 4—figure supplement 2C). OE of Opa1 at 25 °C on its own did not affect contractility (Figure 4M), although the hearts were somewhat constricted (Figure 4—figure supplement 1J and K); however, combining Opa1 OE and Chchd3/6 KD contractility and F-actin staining were further reduced than Chchd3/6 KD alone (Figure 4M, Figure 4—figure supplement 2D), although this interaction did not reach significance (Figure 4—figure supplement 1L). In summary, based on heart contractility and F-actin staining results, we found significant genetic interactions between mitochondrial fission-fusion genes and Chchd3/6, thus supporting the idea that compromised MICOS complex genes affect mitochondrial fission/fusion mechanisms.
Mitochondrial ATP synthase (complex V) KD causes contractile dysfunction and diminished sarcomeric F-Actin staining similar to Chchd3/6 KD
To determine whether KD of other mitochondrial genes impaired contractility and sarcomeric F-actin accumulation similar to that of Chchd3/6 KD, we screened RNAi lines from different mitochondrial functional groups (FlyBase.org GO term mitochondrion: 0005739) using the Hand4.2-Gal-4; tdtK driver for high-throughput heart imaging analysis (see Materials and methods; Vogler, 2021a). Cardiac KD of 17 of the 21 mitochondrial genes tested displayed reduced fractional shortening, most commonly due to systolic dysfunction. However, only KD of Opa1 and ATP synthase subunits reduced both fractional shortening and F-actin staining (Figure 5, Figure 5—figure supplement 1A). Remarkably, F-actin staining in Hand4.2-Gal4, tdtK >ATPsynβ/BRNAi hearts resembled that of Chchd3/6 KD, that is weakly stained myocardial myofibrils relative to ostial myofibrils (Figure 5E). These findings suggested that the heart dysfunction observed upon Chchd3/6 KD may be mediated via defects in ATP synthase.
Figure 5 with 1 supplement see all
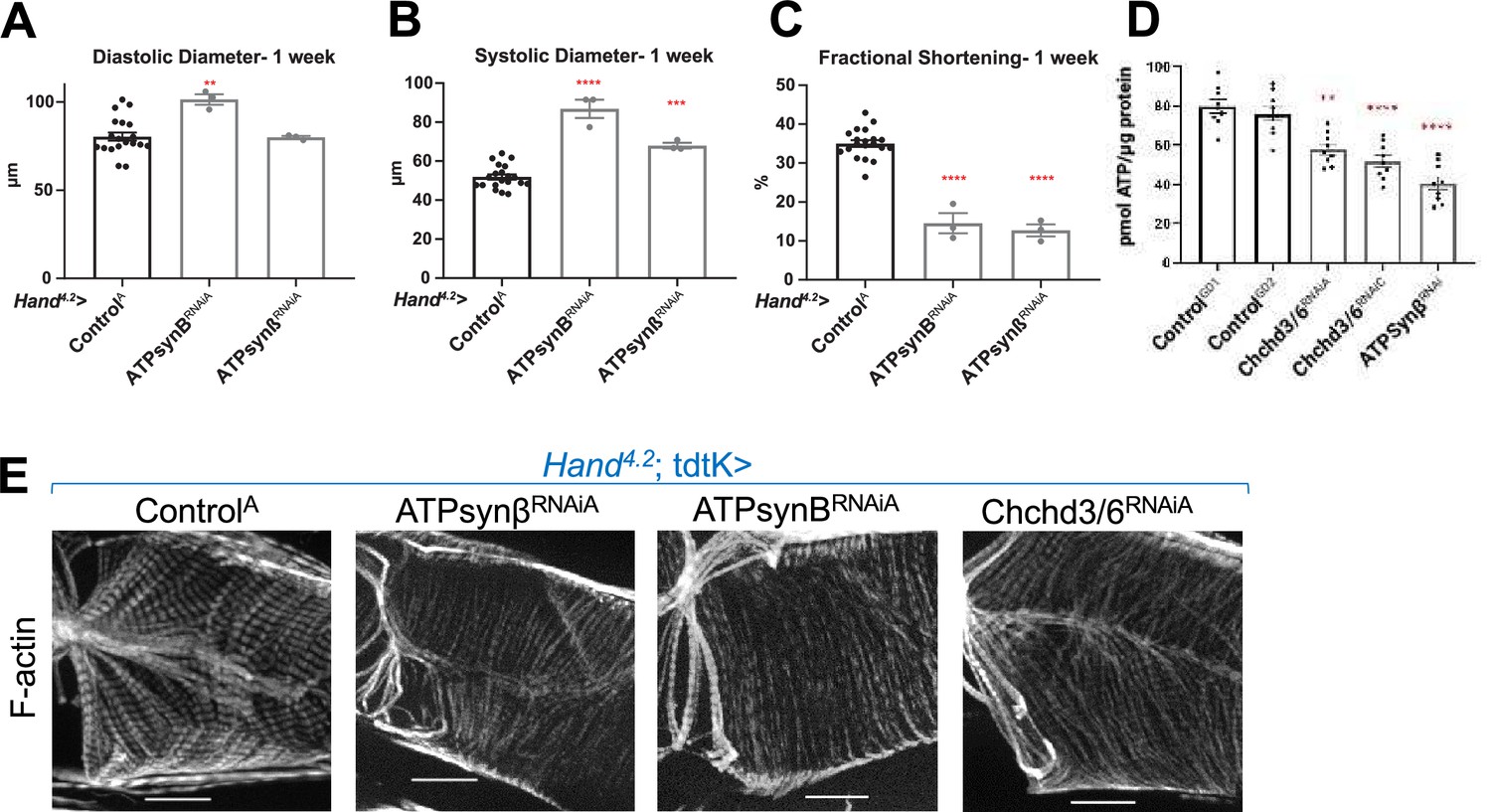
KD of ATP synthase subunit B and beta reduced both fractional shortening and F-actin staining.
(A–C)Hand4.2-Gal4; tdtK driven KD of ATP synthase subunits at 1 week of age measuring (A) diastolic diameter, (B) systolic diameter, and (C) fractional shortening. Data is plotted as ± SEM and significance indicated relative to ControlGD2. ****p ≤ 0.0001, **p ≤ 0.01. (D) Quantification of ATP levels from hearts of 1-week-old flies (10–12 hearts per sample). ATP measurements were plotted relative to protein content. (E) One-week-old Hand4.2-Gal4; tdtK driven KD of ATP synthase subunits with altered F-actin (Chchd3/6 KD is depicted to contrast the structural phenotypes). Statistical differences were calculated by one-way ANOVA followed by Tukey’s post hoc test for multiple comparisons.
ATP production is reduced upon Chchd3/6 knockdown
Disrupted mitochondrial organization and reduced staining of OXPHOS components likely impacts ATP production and could explain cardiac functional and structural defects. We therefore directly measured ATP concentration in 1-week-old female hearts with cardiac Chchd3/6 KD. We observed reduced ATP levels upon Chchd3/6 KD compared to controls, and these ATP levels were similar to those measured in response to cardiac ATP-synβ KD (Figure 5D). This further strengthens the hypothesis that the cardiac functional deficits in contractility induced by Chchd3/6 KD are due to mitochondrial defects that considerably reduce ATP levels in the heart necessary to build and maintain myofibrils.
Chchd3/6 knockdown in all muscle cells is lethal or reduces climbing ability
To further characterize the impact of Chchd3/6 KD induced mitochondrial defects on muscle function, we assessed locomotive ability. When reared throughout development at 25 °C, Mito::GFP; Mef2 >Chchd3/6RNAiA flies were pupal lethal. However, with a moderate strength RNAi line (Mito::GFP; Mef2 >CHCHD3/6RNAiC1) flies did eclose, but with reduced viability, especially for males (Figure 5—figure supplement 1B). Flies are negatively geotactic and will rapidly climb up the sides of a vial when tapped down. In 1 week of age, male and female Mito::GFP; Mef2 >Chchd3/6RNAiC1 flies, this activity was greatly reduced compared to controls (Figure 5—figure supplement 1C), supporting the hypothesis that Chchd3/6 KD reduces muscle function.
Knockdown of SAMM50 ortholog and Mitofilin causes cardiac defects, and SAMM50 genetically interacts with CHCHD3/6
To further explore the role of the MICOS complex to maintain myofibrillar structure, we tested five MICOS complex-associated components for their requirement in cardiac contractility and sarcomeric F-actin levels (Figure 6A). Hand4.2-Gal4-mediated KD of IMMT (Mitofilin) or SAMM50 (CG7639) resulted in a significant reduction in fractional shortening, due to systolic dysfunction, mimicking Chchd3/6 KD defect in contractility, whereas orthologs of APOOL (Mic26/27), MICOS13 (QIL1), or MICOS10 (CG12479) KD did not exhibit significant effects (Figure 6B; Figure 6—figure supplement 1A,B). Of note, none of these five MICOS-associated components displayed detectable reduction in sarcomeric F-actin staining upon KD (Figure 6—figure supplement 1C).
Figure 6 with 2 supplements see all
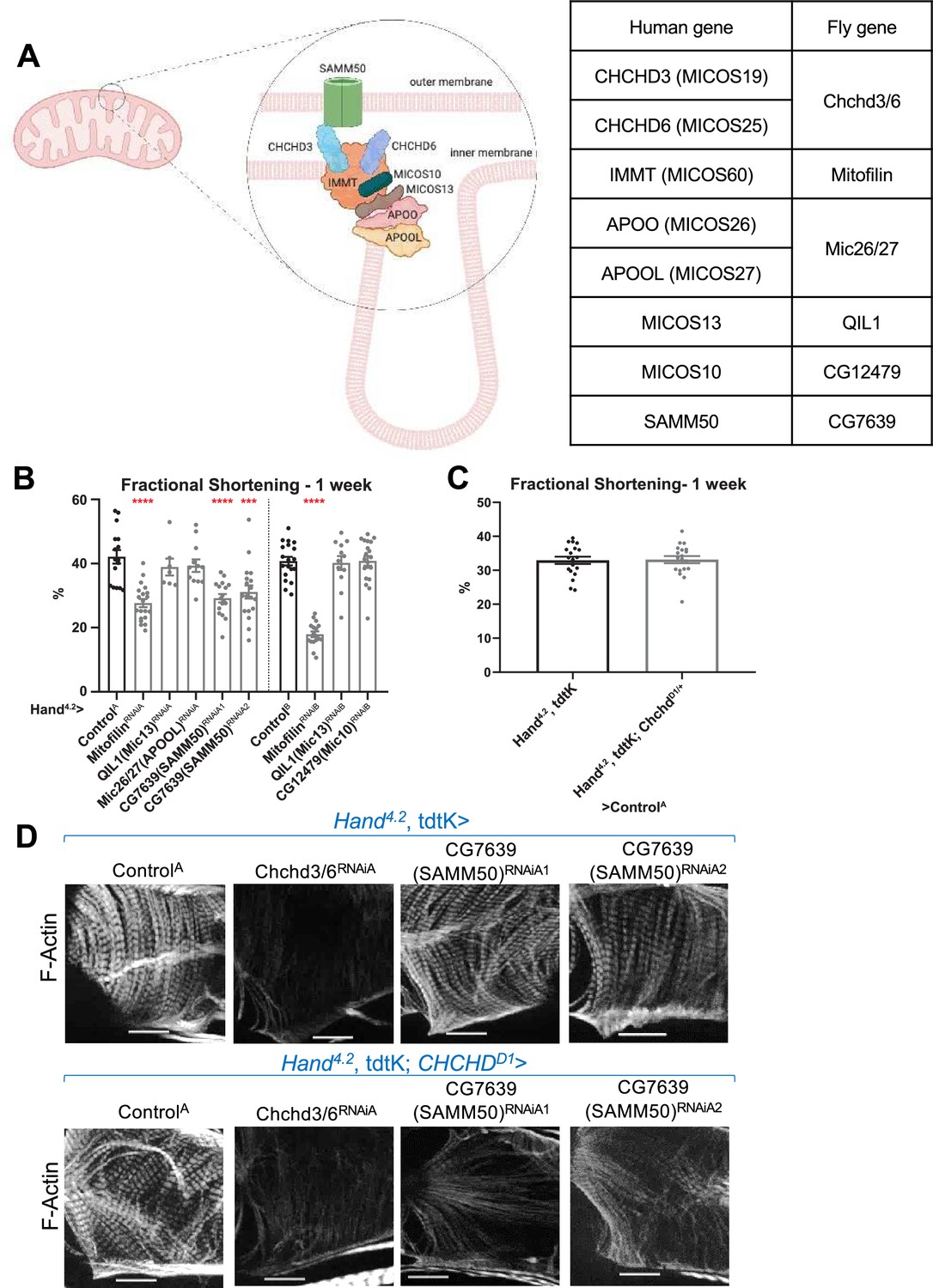
Assessment of other MICOS subunits in the Drosophila heart.
(A) Schematic of the MICOS complex and SAM50. Human MICOS subunits and their respective Drosophila homologs are listed (image created with BioRender.com). (B) Fractional shortening measured from 1-week-old female flies with KD of individual MICOS subunits and Sam50 using a Hand4.2-Gal4 driver. Unpaired two-tailed t-test, ***p≤0.001, ****p≤0.0001; error bars represent SEM. (C) Hand4.2-Gal4, tdtK; ChchdD1/+ line was crossed out with ControlA. Unpaired two-tailed t-test, error bars represent SEM. (D) 1 week old F-actin-stained Drosophila hearts with or without heterozygous loss-of-function ChchdD1/+ in the background, 20 µm scale.
-
Figure 6—source data 1
Human MICOS subunits and their respective Drosophila homologs.
- https://cdn.elifesciences.org/articles/83385/elife-83385-fig6-data1-v2.xlsx
Subsequently, we tested for genetic interactions between Chchd3/6 and MICOS-associated components, since protein-protein interactions among them have been previously described (Ding et al., 2015; Li et al., 2016; Tang et al., 2020). To test for interactions, we generated a Hand4.2-Gal4, tdtK; ChchdD1/+ heterozygote mutant sensitizer line that had no noticeable cardiac abnormalities (Deng et al., 2016; Figure 6C). Hand4.2-Gal4, tdtK; ChchdD1/+ crossed to Mitofilin or CG7639 (SAMM50) RNAi did not further reduce fractional shortening beyond what was observed in response to KD of the individual genes (Figure 6—figure supplement 1D). However, we observed an interaction in the combined SAMM50 KD and ChchdD1/+ hearts, where F-actin levels were also strikingly reduced compared to the single KD, similar the Chchd3/6 KD (Figure 6D). This suggests that there is a threshold requirement of MICOS/SAMM50, which when reached induces reduced contractility AND diminished F-Actin levels.
Knockdown of MICOS subunits impairs proliferation and oxygen consumption of human iPSC-derived cardiomyocytes
We next tested the effects of KD of CHCHD3/6 and other MICOS-associated components in human cardiomyocytes (CMs) derived from iPSC (hiPSC-CM; Cunningham et al., 2017, Yu et al., 2018). Since reduced CM proliferation is hypothesized to be a major contributing factor for the etiology of HLHS (Gaber et al., 2013; Liu et al., 2017; Theis et al., 2020), we focused on proliferation as our readout. We used small interfering RNA (siRNA) (as in Theis et al., 2020) to KD genes in hiPSC-CMs. We found that KD of CHCHD6 and CHCHD3, as well as all other MICOS subunits and SAMM50, significantly reduced their proliferation in an EdU incorporation assay (Figure 6—figure supplement 2A,B), supporting a potential link for CHCHD6 and other MICOS subunits in HLHS pathogenesis.
Moreover, oxygen consumption rate (OCR), was significantly decreased in the combined KD of CHCHD3 and CHCHD6 in hiPSC-CMs 60 min after inhibition of ATP synthase by oligomycin treatment (Figure 6—figure supplement 2C). Similar to the fly heart results, staining of sarcomeric F-actin along the fiber (yellow line in Supp. ) was reduced by co-KD of CHCHD3 and CHCHD6 (Figure 6—figure supplement 2E).
Testing of candidate genes prioritized in HLHS probands with CHCHD3 or CHCHD6 variants reveals novel genetic interactors
Since we found an essential role for Chchd3/6 in establishing heart structure and function in the Drosophila heart model with possible relevance for HLHS pathology, we assessed the presence of variants in additional HLHS family trios. Among the 183 Mayo Clinic HLHS family trios and pediatric cardiac genomics consortium (PCGC) databank (Jin et al., 2017), there were three probands with variants in CHCHD6 (including 11 H) and four with CHCHD3 variants. In total, there were four noncoding variants with a RegulomeDB rank of 2 a (n=2), 2b (n=1) and 4 (n=1) providing evidence for haploinsufficiency due to predicted disruption of transcription factor binding in the presence of the variant. The remaining two missense variants, one inherited and one de novo, may alter the protein structure or function and lead to downstream functional consequences.
The relative abundance of rare, predicted damaging CHCHD3/6 variants in the Mayo Clinic cohort, together with the postulated oligogenic nature of HLHS, led us to test for genetic interactions between CHCHD3/6 and other HLHS candidate genes. Specifically, we prioritized candidate genes with rare coding and regulatory variants identified in HLHS probands who also carried CHCHD3- or CHCHD6-variants. With the Chchd3/6 sensitizer line, Hand4.2-Gal4, tdtK; Chchd3/6RNAiC1, which at 21 °C does not exhibit significant contractility deficits (Figure 7A), we screened 120 RNAi lines representing 60 candidate HLHS genes for genetic interactions and identified three hits that had contractility defects only when co-knocked down with Chchd3/6RNAiC1 at 21 °C (Figure 7B). Cdk12 (human ortholog: CDK12) has been shown to activate RNA polymerase II to regulate transcription elongation (Bartkowiak et al., 2010), goliath (human ortholog: RNF149) encodes an E3 ubiquitin ligase that localizes to endosomes (Yamazaki et al., 2013) and β-Spectrin (human ortholog: SPTBN1) is a scaffolding protein that links the actin cytoskeleton to the plasma membrane. KD of Cdk12 in combination with Chchd3/6 KD also led to greater lethality of elcosed flies at 1 week-of-age compared to individual gene KD (Figure 7C). While interestingly, we found that co-KD of β-Spectrin and Chchd3/6RNAiC1 at 21 °C was the only combination that also diminished F-actin and Myosin staining, similar to co-Chchd3/6RNAiA KD (Figure 7D). In summary, our approach using a sensitized screening strategy to interrogate genetic interactions between patient-specific candidate genes is a powerful tool to identify novel interactions that are potentially involved in the oligogenic ethology of HLHS and other CHDs.
Figure 7
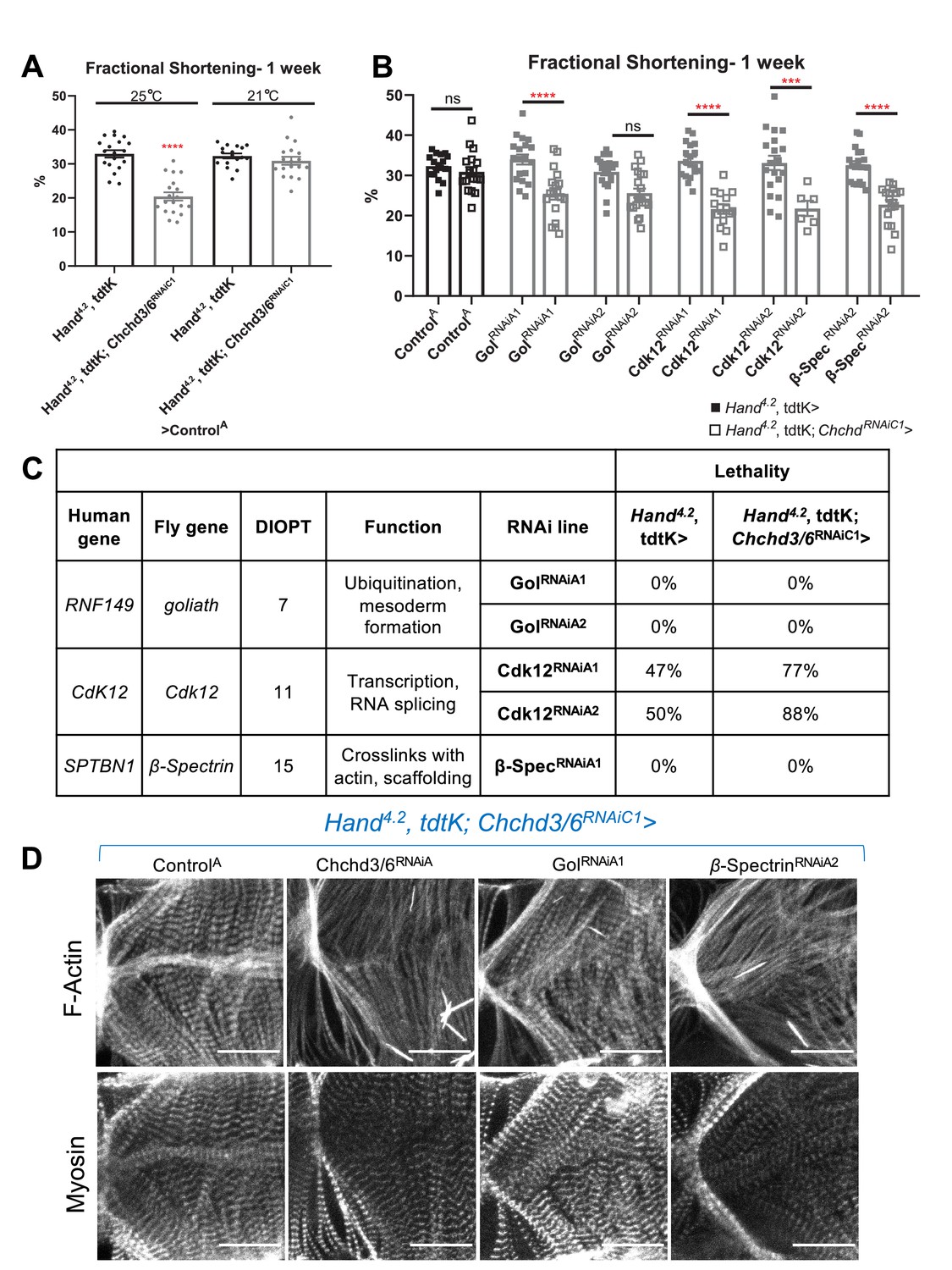
HLHS CHCHD3 and CHCHD6 family-based gene interaction screen reveals three hits.
(A) A Hand4.2-Gal4, tdtK; Chchd3/6RNAiC1 sensitizer line show reduced fractional shortening at 25ºC, which is no longer significant at 21ºC. Unpaired two-tailed t-test, ****p≤ 0.0001; error bars represent SEM. (B) Genetic interaction of Chchd3/6 and prioritized HLHS candidates. Two-way ANOVA with Tukey’s multiple comparisons test, only statistical comparisons between the same RNAi lines are shown; ***p≤ 0.001, ****p≤ 0.0001; error bars represent SEM. (C) Functional overview of human and Drosophila orthologs. KD of Ckd12 with Chchd3/6 KD led to increased lethality of eclosed flies by 1 week-of-age.
-
Figure 7—source data 1
Functional overview of human and Drosophila orthologs.
KD of Ckd12 with Chchd3/6 KD led to increased lethality of eclosed flies by 1 week-of-age.
- https://cdn.elifesciences.org/articles/83385/elife-83385-fig7-data1-v2.xlsx
Discussion
HLHS is characterized by a small left heart, including reduced left ventricle size and mitral and/or atrial atresia or stenosis, and aortic hypoplasia, collectively obstructing systemic blood flow (Tchervenkov et al., 2006). As a consequence, newborns cannot sustain systemic blood flow for more than a few days and therefore require treatment soon after birth. There is a need for improved therapies to treat HLHS patients, and this requires a better understanding of the biology behind HLHS pathogenesis. Here, we probed the genetic basis of HLHS using WGS and powerful bioinformatic gene variant prioritization in a large cohort of HLHS proband-parent trios combined with model system validation.
The 11 H family was prioritized because of consanguinity, implicating a homozygous recessive mode of inheritance that resulted in a short list of nine candidate genes. These candidate genes were probed in Drosophila and iPSC-CMs for a potential role in cardiomyocyte development and function, to gain new insights into HLHS and CHDs in general. Among these HLHS gene candidates, we focused on CHCHD3/6, which has not been previously studied in the heart, and which had striking cardiac functional and structural defects in Drosophila. Specifically, the preliminary gene screen demonstrated that Chchd3/6 cardiac-specific KD caused reduced contractility and decreased sarcomeric F-Actin and Myosin staining.
Our data suggest that Chchd3/6 is necessary during larval and early adult stages to maintain contractility in the adult heart. This is relevant since patients with HLHS have both structural heart disease and risk for later myocardial failure (Theis et al., 2015b). The prevailing ‘no flow, no grow’ hypothesis for HLHS pathogenesis surmises that reduced blood flow in the fetal heart causes underdevelopment of the left ventricle (Goldberg and Rychik, 2016; Grossfeld et al., 2019). A reduced ability for the heart to contract in utero, due to reduced CHCHD6 activity, could contribute to decreased ventricular blood flow in the embryo, resulting in an abnormally small left ventricle. Moreover, reduced CHCHD6 activity could compromise right ventricular function later in life (Theis et al., 2015a; Theis et al., 2015b). In fact, the 11 H proband exhibited mildly reduced right ventricular ejection fraction several years after successful surgical palliation. Consistent with our model system, CHCHD6 deficiency could result in cumulative impairment of mitochondrial function, leading to contractile dysfunction (Sun et al., 2016). Why a mitochondrial defect would have a preferential effect on the left ventricle is still an enigma. We speculate that some of the patient-specific variants that potentially contribute to this likely polygenic disease are in genes that may have a higher expression level or functional importance in the left ventricle, thus in combination with MICOS variants preferentially affecting left-ventricular growth and differentiation, leading to decreased contractility, then again compounded by impaired blood flow feeding back to diminishing growth. Future studies investigating the polygenic basis of HLHS are needed to address this question.
Chchd3/6 KD in the fly heart led to mitochondrial fission-fusion defects, with reduced ATP synthase (complex V) levels, and consequently impaired ATP production. It has previously been reported that CHCHD3 KD in HeLa cells resulted in fragmented mitochondria that was due to improper mitochondrial fusion (Darshi et al., 2011). It has also been demonstrated in yeast that individual or combinatorial loss of MICOS complex proteins disrupt cristae morphology (Friedman et al., 2015), thus suggesting a mechanism by which CHCHD3/6 loss could mediate HLHS pathogenesis. Furthermore, we identified a genetic interaction between SAMM50 and CHCHD3/6 that leads to a contractile deficit and diminished sarcomeric F-Actin. Recent findings demonstrate that SAMM50 directly interacts mammalian CHCHD3, to mediate inner and outer membrane bridging and cristae morphology (Tang et al., 2020).
Our data further suggest that ETC Complex V/ ATP synthase is a potential downstream effector of CHCHD3/6 and MICOS complex function. Individual KD of ATP synthase subunits resulted in reduced fractional shortening and reduced sarcomeric actin. As a result, we hypothesize reduced CHCHD3/6 expression affects ETC function, specifically ATP synthase, leading to reduced ATP production. Since the MICOS complex is in involved ETC assembly in cristae, ATPase subunits may not be assembled correctly causing mitochondrial dysfunction, accompanied by reduced/abnormal mito-GFP staining (see Figure 4B and H). OXPHOS complex assembly has been shown to be disrupted upon MICOS depletion, and we speculate ATP synthase function may be disrupted when CHCHD3/6 is reduced (Cogliati et al., 2016). Consistent with this, we observe depletion of ATP synthase levels upon Chchd3/6 KD.
Finally, we tested a potential oligogenic basis of HLHS in our family-based CHCHD3 and CHCHD6 interaction screen and identified three hits that reduced fractional shortening only in conjunction with CHCHD3/6, but not on their own. Co-KD of Cdk12 and Chchd3/6 also reduced fractional shortening, and caused greater lethality relative to Cdk12 KD alone. Cdk12 activates RNA polymerase II to regulate transcription elongation (Bartkowiak et al., 2010). We postulate that since Chchd3/6 is a nuclear-encoded gene, reducing transcription with Cdk12 KD could decrease CHCHD3/6 levels in a background where CHCHD3/6 activity is already compromised. Alternatively, reduced transcription of other nuclear genes associated with ATP production in combination with Chchd3/6 KD could further reduce ATP levels enough to cause contractility defects. In support of this, a study examining the effects of RMP (RNA polymerase II subunit 5-mediating protein) found that mice with cardiac-specific Rpm KO exhibited reduced fractional shortening and ATP levels, which were attributed to a reduction in mRNA and protein levels of the mitochondrial biogenesis factor PGC1α (Zhang et al., 2019). The second hit, goliath, is an endosomal ubiquitin E3 ligase. Although goliath has been implicated in endosomal recycling (Yamazaki et al., 2013), its role in Drosophila mitophagy in vivo has not been examined. Reduced cardiac contractility with co-KD of gol and Chchd3/6 could result from impaired mitophagy and reduced mitochondrial biogenesis. Together, the accumulation of damaged mitochondria can reduce ATP content required for contraction (Palikaras and Tavernarakis, 2014; Palikaras et al., 2015). The third hit, β-Spectrin, acts as a scaffolding protein. Recent data suggests that the human ortholog, SPTBN1 (Nonerythroid spectrin β) influences SPTAN1 (Nonerythroid spectrin α) levels, which has a calmodulin binding domain (Ackermann and Brieger, 2019). Therefore, decreased β-Spec expression could reduce Calmodulin levels, thereby reducing contractility due to the combined reduction in Ca2+ handling and Chchd3/6 KD-induced reduced ATP levels.
In summary, we have identified a novel mechanism potentially involved HLHS pathogenesis, starting by analyzing WGS data from a prioritized family and large cohort of HLHS patients, followed by functional testing in vivo using the Drosophila heart model and in vitro using human iPSC-derived CMs. Compromised contractile capacity, diminished sarcomeric F-Actin and Myosin accumulation, and mitochondrial dysfunction in Chchd3/6 KD Drosophila hearts are promising phenotypes that could contribute to early HLHS manifestations or heart failure complications later in life. Further examination of the interactions between the MICOS complex and other emerging candidate genes will identify novel gene functions and pathways that contribute to HLHS pathogenesis. Furthermore, a detailed elucidation of novel candidate genes and genetic interactions based on patient-specific rare potentially damaging variants is expected to lead to gene networks that are relevant for HLHS and other CHDs.
Materials and methods
Study subjects
Request a detailed protocolWritten informed consent was obtained for the index family and an HLHS cohort, under a research protocol approved by the Mayo Clinic Institutional Review Board (‘Genetic Investigations in Hypoplastic Left Heart Syndrome’, IRB #11–000114). Participants consented to providing clinical health record data, sample procurement for DNA analyses, and publication of de-identified research findings. Cardiac anatomy was assessed by echocardiography.
Comparative genomic hybridization
Request a detailed protocolTo detect aneuploidy, array comparative genomic hybridization was performed using a custom 180 K oligonucleotide microarray (Agilent, Santa Clara, CA), with a genome-wide functional resolution of approximately 100 kilobases. Deletions larger than 200 kilobases and duplications larger than 500 kilobases were considered clinically relevant.
Genomic and bioinformatics analysis of 11H family
Request a detailed protocolGenomic DNA was isolated from peripheral white blood cells or saliva. WGS and variant call annotation were performed utilizing the Mayo Clinic Medical Genome Facility and Bioinformatics Core. For the family quintet, 101 base pair (bp) or 150 bp paired-end sequencing was carried out on Illumina’s HiSeq 2000 or HiSeq 4000 platforms, respectively. Reads were aligned to the hg38 reference genome using BWA version 0.7.10 (http://bio-bwa.sourceforge.net/bwa.shtml) and duplicate reads were marked using Picard (http://picard.sourceforge.net). Local realignment of INDELs and base quality score recalibration were then performed using the Genome Analysis Toolkit version 3.4–46 (GATK) (McKenna et al., 2010). SNVs and INDELs were called across all samples simultaneously using GATK’s Unified Genotype with variant quality score recalibration (VQSR) (Poplin et al., 2018).
Variant call format (VCF) files with SNV and INDEL calls from each family member were uploaded and analyzed using Ingenuity Variant Analysis software (QIAGEN, Redwood City, CA) where variants were functionally annotated and filtered by an iterative process. Annotated variants were subject to quality filters and required to pass Variant Quality Score Recalibration (VQSR) and have a genotype quality score ≥20. Variants were excluded if they were located in a simple repeat region identified using tandem repeats finder (Benson, 1999) or were found to have a minor allele frequency >1% in gnomAD v2.1 (Karczewski et al., 2020). Second, functional variants were selected, defined as those that impacted a protein sequence, canonical splice site, microRNA coding sequence/binding site, or transcription factor binding site within a promoter validated by ENCODE chromatin immunoprecipitation experiments (Raney et al., 2014). Third, using parental and sibling WGS data, rare, functional variants were then filtered for those that were homozygous recessive in the proband.
Analysis of HLHS cohort for variants in the MICOS complex
Request a detailed protocolWGS was performed on samples from 183 individuals with HLHS and 496 family members by the Mayo Clinic Medical Genome Facility or Discovery Life Sciences. SNVs and INDELs that passed quality control were subject to further filtering based upon rarity (MAF <0.01) and predicted consequence. Details about the sequencing and subsequent variant filtering to identify HLHS gene candidates have been previously described (Theis, 2021). Rare variants from the 183 probands were interrogated for variants in CHCHD3 and CHCHD6 to identify variants that arose de novo or were homozygous recessive, compound heterozygous or X-linked recessive. Next, inherited variants in these genes were analyzed, but stricter thresholds were required to identify the most damaging variants. Missense variants were required to have CADD >24 (corresponds to the upper quartile of the most damaging missense variants) and non-coding variants were required to have a Position Weight Matrix (PWM) score >0.75 from the Factorbook database (selecting for variants predicted to disrupt canonical transcription factor binding sites). In addition to the Mayo Clinic HLHS cohort, the Pediatric Cardiac Genomics Consortium (PCGC) whole exome sequencing dataset was interrogated for candidate genes in the MICOS complex in patients with CHD (Jin et al., 2017).
Analysis of CHCHD3 and CHCHD6 variant carriers
Request a detailed protocolUsing robust bioinformatics algorithms as previously described (Theis and Olson, 2022), a broad range of both family-based Mendelian inheritance modeling and cohort-wide enrichment analyses were applied to identify additional candidate genes in HLHS probands identified to have a rare, predicted-damaging coding or regulatory variant in CHCHD3 or CHCHD6.
Drosophila strains and husbandry
Request a detailed protocolDrosophila crosses were reared and aged at 25 °C, unless otherwise noted. Drosophila orthologs were determined using DIOPT (Drosophila RNAi Screening Center Integrative Ortholog Prediction Tool) which calculates the number of databases that predict orthology (out of a score of 16) (Hu et al., 2011). Fly stocks were obtained from Vienna Drosophila Resource Center (VDRC) and Bloomington Drosophila Stock Center (BDSC). Lines include Hand4.2-Gal4 (Han et al., 2006), tdtK (Klassen et al., 2017), tinCΔ4-Gal4 (Lo and Frasch, 2001), Dot-Gal4 (Kimbrell et al., 2002), Mef2-Gal4 (Ranganayakulu et al., 1998), Mito::GFP (BDSC: 8442), CHCHDDefA (BL: 26847), CHCHD3/6RNAiA (VDRC: 52251), CHCHD3/6RNAiB (VDRC: 105329), CHCHD3/6RNAiC1 (BDSC: 51157), CHCHD3/6RNAiC2 (BDSC: 38984), DuoxRNAi (BDSC: 32903), MitofilinRNAiA (VDRC: 47615), MitofilinRNAiB (VDRC: 106757), QIL1(Mic13)RNAiA (VDRC: 14283), QIL1(Mic13)RNAiB (VDRC: 100911), CG12479(Mic10)RNAiA (VDRC: 102479), Mic26/27(APOOL)RNAiA (VDRC: 31098), CG7639(SAMM50)RNAiA (VDRC: 33641), CG7639(SAMM50)RNAiB (VDRC: 33642), Drp1RNAiA (VDRC: 44155), Drp1RNAiC (BDSC: 27682), Drp1OE (BDSC: 51647), Opa1RNAiB (VDRC: 106290), Opa1RNAiC (BDSC: 32358) and Opa1OE (BDSC: 95258). CHCHDD1 was kindly shared by the Ge lab (Deng et al., 2016).
Assessment of lethality in co-KD of CHCHD3/6 and Cdk12 refers to percentage of surviving flies at 1 week-of-age, versus the number of flies eclosing on day 0.
In situ heartbeat analysis
Request a detailed protocolAn in-situ dissection approach was used to expose the denervated beating fly heart (Fink et al., 2009; Ocorr et al., 2009; Vogler and Ocorr, 2009). SOHA (Semi-automated optical heartbeat analysis) was used to analyze high speed video recordings to determine heart-related parameters (Fink et al., 2009). Flies (n>15) were briefly anesthetized using filter paper with 10 µm FlyNap and transferred to a 10X35 mm Petri dish with Vaseline to attach the hydrophobic wing cuticle to the dish. Oxygenated room temperature artificial hemolymph (108 mM NaCl, 5 mM KCl, 2 mM CaCl2•2H2O, 8 mM MgCl2•6H2O, 15 mM pH 7.1 HEPES, 1 mM NaH2PO4•H2O, 4 mM NaHCO3, 10 mM sucrose, and 5 mM trehalose) was added to each dish. Flies were dissected as per Vogler and Ocorr, 2009 and oxygenated for minimum 15 min to equilibrate. Dissected flies were filmed for 30 s using an Olympus BX63 microscope (10 X magnification), a Hamamatsu C11440 ORCA-flash4.0 OLT digital camera, and HCImageLive program. These videos were uploaded to SOHA (semi-automated optical heartbeat analysis), end diastolic and end systolic diameters were manually marked towards end of ostia, and heart-related parameters were extracted (Fink et al., 2009).
In vivo heartbeat analysis
Request a detailed protocolNorland #61 optical glue was placed on a 22X50 mm coverslip (one small drop for each fly). Flies (n>15) were briefly anesthetized using filter paper with 10 µm FlyNap, transferred to coverslip on individual adhesive drops with the dorsal side facing the coverglass, and cured for 30 s using ultraviolet light. The coverslip was then placed on a 10X35 mm Petri dish and secured using putty. Fly hearts were filmed for 5 s using an Olympus BX63 microscope (×20 magnification), a Hamamatsu C11440 ORCA-flash4.0LT digital camera, and HCImage Live program. All analysis was automatically processed using R (Vogler, 2021a).
Adult Drosophila heart immunohistochemistry
Request a detailed protocolFlies were dissected as per Vogler and Ocorr, 2009 in a 10X35 mm Petri dish and EGTA was added to a final concentration of 10 mM. EGTA was removed and replaced with 4% methanol-free formaldehyde for 20 min. Formaldehyde was removed and replaced with 1 X PBS 3 times. Fly thoraxes were removed, abdominal walls were trimmed, and excess fat around heart was removed. Hearts were then washed three times with 0.3% PBTx (Triton-X) for 15 min on a shaker. PBTx was removed and replaced with 200 µL 1° antibody solution (0.3% PBTx +1° antibody), then a small piece of Parafilm with carefully placed over the solution to form and seal of liquid over the hearts. Dishes were incubated either (1) at 4 °C overnight or (2) at room temperature for 2 hr while shaking. Once finished incubating, Parafilm was gently removed with forceps and three 15-min washes with 0.3% PBTx were performed. PBTx was removed and replaced with 200 µL 2° antibody solution (0.3% PBTx +2° antibody), then a small piece of Parafilm was carefully placed over the solution. Dishes were incubated either (1) at 4 °C overnight or (2) at room temperature for 2 hr while shaking. Parafilm was gently removed with forceps and three 15-min washes with 0.3% PBTx were performed. PBTx was removed and replaced with 1 X PBS. Ventral cuticle with attached hearts were carefully removed individually from the Vaseline layer and transferred to a 25X75 X 1mm slide with 2 18X18 mm No. 1 coverslips glued to form a bridge and ProLong Gold antifade mounting medium (Invitrogen) in the middle. Flies were placed ventral side up and covered with a 18X18 mm No. 1.5 coverslip, sealed with clear nail polish around the edges, and stored at room temperature for 24 hr until being moved to 4 °C.
Primary Antibodies: anti-dMef2 (1:20, gift from Dr. Bruce Paterson); anti-Slit (1:40, c555 DSHB); anti-Myosin (1:50, 3E8-3D3 DSHB); anti-Sallimus (1:100, Abcam); anti-ATP5A (1:100, Abcam 14748); anti-ATP5A1 (1:200, Invitrogen 43–9800). All Secondaries from Jackson Immuno Research Labs used at 1:500: Goat anti-Rat 594; Goat anti-Rabbit 647; Goat anti-Rabbit Cy5; Goat anti-mouse Cy3. Dyes: Phallodin 594 or 647 (1:100, Invitrogen).
Drosophila indirect flight muscle dissection and immunofluorescence
Request a detailed protocolThoraxes were removed under light CO2 pressure and fixed for 40 min in 5% PFA, followed by three 2-min PBS washes. IFM muscle fibers were removed using fine (#55) forceps and washed with 0.5% PBTx for 15 min, then washed with 0.1% PBTx twice for 15 min. All subsequent antibody stainings were diluted in 0.1% PBTx and incubated shaking at 4 °C overnight to penetrate the muscle tissue. IFMs were transferred to a 25X75 X 1mm slide without a bridge. ProLong Gold antifade mounting medium (Invitrogen) was added, the samples were covered with an 18X18 mm No. 1.5 coverslip, and sealed with clear nail polish around the edges.
Drosophila embryo collection, fixation, and immunofluorescence
Request a detailed protocolAdult flies were reared in a plastic bottle cage with a Petri dish on the bottom containing grape agar (Agar, EtOH, glacial acetic acid, grape juice) and yeast paste (yeast and H2O) at 25 °C. After incubating (16 hr), embryos were carefully collected with a brush and placed in a mesh basket. Flies were washed with water, then placed in bleach for 3 min, followed by 30 s of wash with water. Embryos were removed from mesh and placed in a fixation solution (2 Heptane: 1 2 X PBS: 1 10% formaldehyde) for 25 min. Formaldehyde layer (bottom) was removed, replaced with 500 µL MetOH, vortexed, then the supernatant with vitelline membranes (middle layer) was removed, this was repeated once more. Embryos were washed with MetOH (3 rinses, followed by 1 hr on rotator). Embryos were stored in fresh MetOH at –20 °C. 1° antibody was added and tube was placed on rotator at 4 °C overnight. 1° was later removed using 3X15 min 0.4% PBTx washes. PBTx was removed, replaced with 2° antibody (diluted in PBTx), and rotated for 2 hr. 2° was later removed using 3X15 min 0.4% PBTx washes, then left in PBS at 4 °C. Since ChchdD1 and ChchdDefA are both homozygous lethal at adult stages, each line was rebalanced over TM6b YFP (ChchdD1/TM6b YFP and ChchdDefA/TM6b YFP). The YFP lines (ChchdD1/TM6b YFP and ChchdDefA/TM6b YFP) were crossed out and embryos were selected against GFP to obtain only ChchdD1/ChchdDefA embryos.
Fixed sample imaging
Request a detailed protocolSamples were imaged at ×10, ×25, or ×40 magnification using a Zeiss Apotome.2 Imager Z1, a Hamamatsu C11440 ORCA-Flash4.0LT digital camera, and Zeiss ZEN. In order to obtain higher resolution, confocal microscopy was performed for all immunohistochemistry experiments involving Mito::GFP and ATP synthase staining.
Climbing assay
Request a detailed protocolFlies were initially anesthetized using FlyNap, placed into five separate vials, and counted for a total at week 0 for Control C females = 178, Control C males = 125, Chchd3/6 C females = 154, Chchd3/6 C males = 112. Each week, flies were transferred using a funnel to a clean longer tube with no food and these tubes were placed in a Styrofoam cutout to hold the tubes for tapping. The vial holder was tapped down multiple times until flies were at the base of the vial and then left to record the percentage of flies which reached 10 cm after 10 s. The vial holder was tapped down multiple time to achieve biological replicates = 4 and different batches of genotypes were examined for technical replicates = 5.
Statistical analyses
Request a detailed protocolAll statistical analyses were performed using GraphPad Prism version 8.0.1 for Windows, GraphPad Software, San Diego, California USA, https://www.graphpad.com. Statistical tests used are stated in figure legends. T-tests were performed on most heart assays where only one variable was defined. For tdtK analyses, a ranked one-way ANOVA Kruskal–Wallis test was used. Combinatorial KD assays with MICOS subunits or mitochondrial fission-fusion genes and Chchd3/6 loss-of-function in Drosophila were analyzed with two-way ANOVA. The interaction plots of two-way ANOVA in Figure 4—figure supplement 1 were obtained using the “Plot2WayANOVA” function in the “CGPfunctions” package in R.
PCR
Request a detailed protocolThe ChchdD1 line was confirmed via PCR using primers ChchdD1 F4: ATATATCCGACGATGTGG and ChchdD1 R4: AGCTCCTGGTTCATCTGG (Q5 High-Fidelity 2 X Master Mix New England Bio).
Quantitative real time polymerase chain reaction (qRT-PCR)
Request a detailed protocolRNA was extracted using Qiagen miRNeasy Mini Kit and cDNA was synthesized with Qiagen QuantiTech reverse transcription kit. qRT-PCR analysis was performed using Roche FastStart Essential DNA Probes Master and Roche LightCycler 96 with 2 biological replicates and 3 technical replicates. Data was analyzed in the LightCycler application. Primers include Chchd3/6 F: GCTAGAGGAACTTCAAAGATGG, Chchd3/6 R: GGGATAGGAGGATACTTTCGG, RP49 F: GCTAAGCTGTCGCACAAATG, RP49 R: GTTCGATCCGTAACCGATGT.
Human iPSC-derived cardiomyocyte proliferation assays
Request a detailed protocolAt day 25 of differentiation, human iPSC-derived cardiomyocytes (hiPSC-CMs) were dissociated with TrypLE Select 10 X (Gibco) for up to 12 min and action of TrypLE was neutralized with RPMI supplemented with 10% FBS. Cells were resuspended in RPMI with 2% KOSR (Gibco) and 2% B27 50 X with vitamin A (Life Technologies) supplemented with 2 µM Thiazovivin and plated at a density of 5000 cells per well in a Matrigel-coated 384-well plate. hiPSC-CMs were then transfected with siRNA (Dharmacon) directed against each gene using lipofectamine RNAi Max (Thermo Fisher). Each siRNA was tested in quadruplicate. Forty-eight hours post-transfection, cells were labeled with 10 µM EdU (Thermo Fisher). After 24 hr of EdU incubation, cells were fixed with 4% paraformaldehyde for 30 min. EdU was detected according to the protocol and cells were stained with cardiac specific marker ACTN2 (Sigma A7811, dilution 1:800) and DAPI. Cells were imaged with ImageXpress Micro XLS microscope (Molecular Devices) and custom algorithms were used to quantify EdU +hiPSC CMs. Cell lines were checked for mycoplasma contamination and genotypic authenticity.
ATP Measurements
Request a detailed protocolMeasurements of ATP were performed using a luciferase assay as described previously (Liu and Lu, 2010; Zanon et al., 2017). 10–12 hearts per sample were collected from 1-week-old flies and homogenized in 100 µl extraction buffer (100 mM Tris and 4 mM EDTA, pH 7.8) containing 6 M guanidine-HCl followed by rapid freezing in liquid nitrogen. The samples were boiled for 5 min and cleared by centrifugation at 14,000 x g. Supernatants were diluted 1:50 and ATP levels were determined using ENLITEN ATP Assay System (Promega, Cat #FF2000) as per manufacturer instructions. Total protein levels were determined by BCA method (Pierce, Cat #23225). ATP measurements were normalized to protein.
Hybridization chain reaction (HCR)
Request a detailed protocolHearts were exposed as described above and RNA in situ performed and analyzed as described in Kirkland et al., 2021. Briefly, hearts were relaxed using 10 mM EGTA in artificial hemolymph and fixed in 4% formaldehyde in 0.1% Tween 20-PBS for 20 min. Hearts were then washed with 0.1% Tween 20, PBS, 2x5 min. On ice, hearts were incubated in a methanol gradient with PBS for 5 min each (25%, 50%, 75%, 100%, 75%, 50%, 25%). Hearts were then permeabilized in 1% Triton 100 X in PBS for 2 hr at room temperature. The hearts were post-fixed with 4% formaldehyde in 0.1% Tween 20-PBS for 20 min at room temperature before washing on ice with 0.1% Tween 20-PBS, 2x5 min. Subsequently, samples were washed with 50%–0.1% Tween 20-PBS and 50% 5XSSCT (5 X SSC, 0.1% Tween 20, H2O) for 5 min on ice, followed by 5 X SSCT for another 5 min. The hearts were then transferred to a 96-well plate and the hearts incubated in probe hybridization buffer (Molecular Instruments) for 5 min on ice, then 30 min at 37 °C. The solution was then replaced with 2 µl of each probe in 200 µl of probe hybridization buffer and incubated at 37 °C overnight (up to 16 hr). Next, 4x15 min washes were performed with probe wash buffer (Molecular Instruments) at 37o C, then 2x5 min 5XSSCT and 1x5 min amplification buffer (Molecular Instruments). Two µl of corresponding h1 and h2 hairpins (Molecular Instruments) were heated to 95 °C for 90 s, cooled in the dark for 30 min and added to 100 µl of amplification buffer. Hairpin solution was then incubated with the heart samples at room temperature overnight (up to 16 hr), in the dark. Next, samples were washed 2x5 min with 5 X SSCT; 2x30 min with 5 X SSCT; 1x5 min with 5 X SSCT and rinsed 3 x with PBS. DAPI in PBS (1:500) was incubated with the samples for 15 min, then samples were again rinsed 3x5 min in PBS. Samples were then mounted and imaged as described above. To quantify expression, a maximum projection image was created in ImageJ from the confocal stack image and binarized. The region around the cardiomyocyte nucleus was traced and the ROI copied to the binary image for particle analysis. Since segmentation was imperfect for transcripts very close together and to account for differences in pocket size, the % area covered by the transcripts was used to assess statistical significance in Prism (GraphPad).
Data availability
Data available on Dryad: https://doi.org/10.5061/dryad.z8w9ghxj1.
-
Dryad Digital RepositoryMitochondrial MICOS complex genes, implicated in hypoplastic left heart syndrome, maintain cardiac contractility and actomyosin integrity.https://doi.org/10.5061/dryad.z8w9ghxj1
References
-
The role of Nonerythroid Spectrin ΑII in cancerJournal of Oncology 1:7079604.https://doi.org/10.1155/2019/7079604
-
Conserved signaling mechanisms in Drosophila heart developmentDevelopmental Dynamics 246:641–656.https://doi.org/10.1002/dvdy.24530
-
The Genetics and pathology of mitochondrial diseaseThe Journal of Pathology 241:236–250.https://doi.org/10.1002/path.4809
-
Two-dimensional echocardiographic assessment of right ventricular function as a Predictor of outcome in Hypoplastic left heart syndromeThe American Journal of Cardiology 86:964–968.https://doi.org/10.1016/s0002-9149(00)01131-0
-
Tandem repeats finder: a program to analyze DNA sequencesNucleic Acids Research 27:573–580.https://doi.org/10.1093/nar/27.2.573
-
Dissociation of mitochondrial from Sarcoplasmic reticular stress in Drosophila cardiomyopathy induced by Molecularly distinct mitochondrial fusion defectsJournal of Molecular and Cellular Cardiology 80:71–80.https://doi.org/10.1016/j.yjmcc.2014.12.018
-
Heart development in Drosophila and its relationship to vertebratesTrends in Cardiovascular Medicine 5:21–28.https://doi.org/10.1016/1050-1738(94)00032-Q
-
BookDevelopment and Aging of the Drosophila Heart, Heart Development and RegenerationElsevier Inc.
-
Evidence for an ATP cap at the ends of actin filaments and its regulation of the F-actin steady stateJournal of Biological Chemistry 259:9983–9986.https://doi.org/10.1016/S0021-9258(18)90914-2
-
Control of actin DynamicsCurrent Opinion in Cell Biology 10:45–51.https://doi.org/10.1016/s0955-0674(98)80085-9
-
Mitochondrial cristae: where beauty meets FunctionalityTrends in Biochemical Sciences 41:261–273.https://doi.org/10.1016/j.tibs.2016.01.001
-
Control of cardiac development by an Evolutionarily conserved transcriptional networkDevelopmental Biology 246:14–28.https://doi.org/10.1006/dbio.2002.0666
-
Re-evaluation of Hypoplastic left heart syndrome from a developmental and morphological perspectiveOrphanet Journal of Rare Diseases 12:1–10.https://doi.org/10.1186/s13023-017-0683-4
-
Id genes are essential for early heart formationGenes & Development 31:1325–1338.https://doi.org/10.1101/gad.300400.117
-
Chchd3, an inner mitochondrial membrane protein, is essential for maintaining Crista integrity and mitochondrial functionThe Journal of Biological Chemistry 286:2918–2932.https://doi.org/10.1074/jbc.M110.171975
-
Fetal Reprogramming and Senescence in Hypoplastic left heart syndrome and in human Pluripotent stem cells during cardiac differentiationThe American Journal of Pathology 183:720–734.https://doi.org/10.1016/j.ajpath.2013.05.022
-
BookHypoplastic left heart syndromeIn: Wyman W, Lai MD, Luc L. Mertens MD, Meryl S, Cohen MD, Tal Geva MD, editors. Echocardiography in Pediatric and Congenital Heart Disease: From Fetus to Adult. wiley. pp. 357–381.https://doi.org/10.1002/9781118742440.ch20
-
Hypoplastic left heart syndrome: A new paradigm for an old diseaseJournal of Cardiovascular Development and Disease 6:1–13.https://doi.org/10.3390/jcdd6010010
-
The MICOS complex of human mitochondriaCell and Tissue Research 367:83–93.https://doi.org/10.1007/s00441-016-2433-7
-
Mic60/Mitofilin determines MICOS assembly essential for mitochondrial Dynamics and mtDNA Nucleoid organizationCell Death and Differentiation 23:380–392.https://doi.org/10.1038/cdd.2015.102
-
The complex Genetics of Hypoplastic left heart syndromeNature Genetics 49:1152–1159.https://doi.org/10.1038/ng.3870
-
Strategic approaches to unraveling genetic causes of cardiovascular diseasesCirculation Research 108:1252–1269.https://doi.org/10.1161/CIRCRESAHA.110.236067
-
Notch1 mutations in individuals with left ventricular outflow tract malformations reduce ligand-induced signalingHuman Molecular Genetics 17:2886–2893.https://doi.org/10.1093/hmg/ddn187
-
Hypoplastic left heart syndromePaediatrics and Child Health 27:75–82.https://doi.org/10.1016/j.paed.2016.12.002
-
Semi-automated optical heartbeat analysis of small heartsJournal of Visualized Experiments 31:1435.https://doi.org/10.3791/1435
-
Sam50 functions in mitochondrial Intermembrane space bridging and Biogenesis of respiratory complexesMolecular and Cellular Biology 32:1173–1188.https://doi.org/10.1128/MCB.06388-11
-
Mitochondrial homeostasis: the interplay between Mitophagy and mitochondrial BiogenesisExperimental Gerontology 56:182–188.https://doi.org/10.1016/j.exger.2014.01.021
-
Mitochondrial quality control and muscle mass maintenanceFrontiers in Physiology 6:422.https://doi.org/10.3389/fphys.2015.00422
-
The mitochondrial basis of agingMolecular Cell 61:654–666.https://doi.org/10.1016/j.molcel.2016.01.028
-
The nomenclature, definition and classification of Hypoplastic left heart syndromeCardiology in the Young 16:339–368.https://doi.org/10.1017/S1047951106000291
-
Celsr1 risk Alleles in familial bicuspid aortic valve and Hypoplastic left heart syndromeCirculation. Genomic and Precision Medicine 15:e003523.https://doi.org/10.1161/CIRCGEN.121.003523
-
Whole genome sequencing in Hypoplastic left heart syndromeJournal of Cardiovascular Development and Disease 9:117.https://doi.org/10.3390/jcdd9040117
-
Impact of Myh6 variants in Hypoplastic left heart syndromePhysiological Genomics 48:912–921.https://doi.org/10.1152/physiolgenomics.00091.2016
-
Drosophila tools and assays for the study of human diseasesDisease Models & Mechanisms 9:235–244.https://doi.org/10.1242/dmm.023762
-
Visualizing the beating heart in DrosophilaJournal of Visualized Experiments 31:1425.https://doi.org/10.3791/1425
-
The genetic landscape of Hypoplastic left heart syndromePediatric Cardiology 39:1069–1081.https://doi.org/10.1007/s00246-018-1861-4
-
Generation of first heart field-like cardiac progenitors and ventricular-like cardiomyocytes from human Pluripotent stem cellsJournal of Visualized Experiments 136:57688.https://doi.org/10.3791/57688
Article and author information
Author details
Funding
Foundation for the National Institutes of Health (HL054732)
- Rolf Bodmer
Wanek Foundation (Todd and Karen Wanek Family Program for HLHS)
- Timothy J Nelson
- Jeanne L Theis
- Timothy M Olson
- Rolf Bodmer
- Alexandre R Colas
American Heart Association (18PRE33960593)
- Katja Birker
American Heart Association (20POST35180048)
- Natalie J Kirkland
Foundation for the National Institutes of Health (HL153645)
- Alexandre R Colas
Foundation for the National Institutes of Health (HL148827)
- Alexandre R Colas
Foundation for the National Institutes of Health (HL149992)
- Alexandre R Colas
Foundation for the National Institutes of Health (AG071464)
- Alexandre R Colas
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We gratefully acknowledge the patients and families who participated in this study. We thank Marco Tamayo and Bosco Trinh for excellent technical assistance. This work was supported by National Institutes of Health (R01 HL054732 to R.B.). This work was also supported by a grant from the Wanek Foundation at Mayo Clinic in Rochester, M.N., to J.L.T., T.J.N., T.M.O., R.B. and A.R.C.; by the American Heart Association: AHA Predoctoral Fellowship (18PRE33960593 to K.B.) and AHA Postdoctoral Fellowship (20POST35180048 to N.J.K.).This work was supported by R01 HL153645, R01 HL148827, R01 HL149992, R01 AG071464 (National Institutes of Health to A.R.C). Sanford Burnham Prebys Shared Resources are supported by an NCI Cancer Center Support Grant (P30 CA030199).
Ethics
Human subjects: Written informed consent was obtained for the index family and an HLHS cohort, under a research protocol approved by the Mayo Clinic Institutional Review Board ("Genetic Investigations in Hypoplastic Left Heart Syndrome", IRB #11-000114). Participants consented to providing clinical health record data, sample procurement for DNA analyses, and publication of de-identified research findings.
Copyright
© 2023, Birker, Ge, Kirkland et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,678
- views
-
- 220
- downloads
-
- 18
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 18
- citations for umbrella DOI https://doi.org/10.7554/eLife.83385
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Mitochondrial MICOS complex genes, implicated in hypoplastic left heart syndrome, maintain cardiac contractility and actomyosin integrity
eLife 12:e83385.
https://doi.org/10.7554/eLife.83385




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}