Neural Plasticity: Restoring cerebellar-dependent learning
Behavioral and pharmaceutical interventions reverse defects associated with increased cerebellar long-term depression in a mouse model of Fragile X syndrome.
- Department of Psychology, Arizona State University, United States
Neurodevelopmental disorders, such as Fragile X syndrome, are often characterized by neurons connecting incorrectly, leading to impairments in learning, memory and cognitive flexibility (Schmitt et al., 2022). Yet the brain can reorganize itself in response to an organism’s changing needs by altering the strength and number of connections (or synapses) between certain neurons. Whether therapeutics could restore behavioral deficits by encouraging nerve cells to 'rewire,' particularly when the brain is at its most malleable, remains a controversial question. In fact, enhancing neural plasticity has been shown to sometimes impair learning (Navakkode et al., 2022; Makin and Krakauer, 2023; Diniz and Crestani, 2023).
Long-term depression (LTD for short) is a type of plasticity that results in weaker synapses. Perhaps counterintuitively, this process is crucial to fine-tune neural responses and enable learning, memory and cognitive flexibility. For example, this is the case in the cerebellum, the brain center best known for controlling movement and learning new motor skills. There, output neurons, known as Purkinje cells, process and integrate the information received from parallel and climbing fibers originating from cells in other parts of the cerebellum or the nervous system. Both types of fibers work together to regulate the strength of the synapses between parallel fibers (PF for short) and Purkinje cells via LTD (Ito, 1989).
Oculomotor learning is a cerebellar mechanism by which the brain can use previous experiences to fine-tune how eye muscles compensate for head movements so that vision can remain steady (a process known as the vestibulo-ocular reflex, or VOR; Nguyen-Vu et al., 2017). Mice with increased LTD at the PF-Purkinje cell synapses have disrupted oculomotor learning, but why this is the case has remained unclear. Now, in eLife, Amin Shakhawat, Jennifer Raymond and colleagues at Stanford University report how this defect can be reverted in a mouse model of Fragile X syndrome (Shakhawat et al., 2023).
Scientists often use the VOR to examine cerebellar function and synaptic plasticity, as animals can be trained to adjust the strength of this reflex (Hirano and Inoshita, 2021; Ito, 1989; Clopath et al., 2014). For instance, mice trying to keep track of rotating black and white stripes while moving on a turntable will make smaller eye movements if this stimulus and the head are moving in the same direction (VOR-decrease learning). Conversely, these eye movements will increase if the stripes and the head travel in opposite directions (VOR-increase learning).
VOR-increase training relies heavily on LTD, yet studies in mice show that such learning is impaired when this type of plasticity is enhanced at PF-Purkinje cells; on the other hand, VOR-decrease learning, which is less dependent on long-term depression, is preserved. This led Shakhawat et al. to propose that enhanced LTD can be too easily triggered by erroneous signals; the cerebellar circuit responds by raising the threshold required to activate this plasticity, which then makes it more difficult for LTD to be recruited during learning.
To test this hypothesis, the team used mice in which Purkinje cells carry the mutation responsible for Fragile X syndrome – a genetic change known to enhance PF-Purkinje cell LTD. First, Shakhawat et al. confirmed that these animals (known as L7-Fmr1 KO mice) showed profound impairment in their VOR-increase learning, while their VOR-decrease learning remained intact. Next, they tested whether two types of interventions that could prevent or reduce a prior increase in activation threshold for LTD might be able to reverse these defects.
The behavioral intervention consisted of both wild-type and L7-Fmr1 KO mice receiving a VOR-decrease ‘pre-training’ session (which can improve LTD at PF-Purkinje cell synapses) before their VOR-increase training. This manipulation did not affect the control group, but it greatly improved VOR-increase learning in the mutated mice.
In the pharmaceutical intervention, Shakhawat et al. treated wild type and L7-Fmr1 KO animals with diazepam prior to VOR as this compound is known to decrease neural firing in the cerebellum (and would therefore prevent incorrect LTD activation prior to testing). Similar to pre-training, diazepam temporarily rescued VOR-increase learning impairments in L7-Fmr1 KO mice if administered 18-24hours before the task. The team further showed that diazepam could restore another type of LTD-dependent learning (known as the optokinetic reflex) for which these mice are also impaired. In fact, receiving this treatment led to the animals outperforming their wild-type counterparts.
Taken together, these results suggest that defects in VOR-increase learning can be rescued through specific pre-training or the use of certain pharmaceuticals (Figure 1). The findings support a model in which prior experiences reshape the activation thresholds of mice with enhanced LTD at their PF-Purkinje cells in such a way that subsequent activation of this plasticity is hindered.
Figure 1
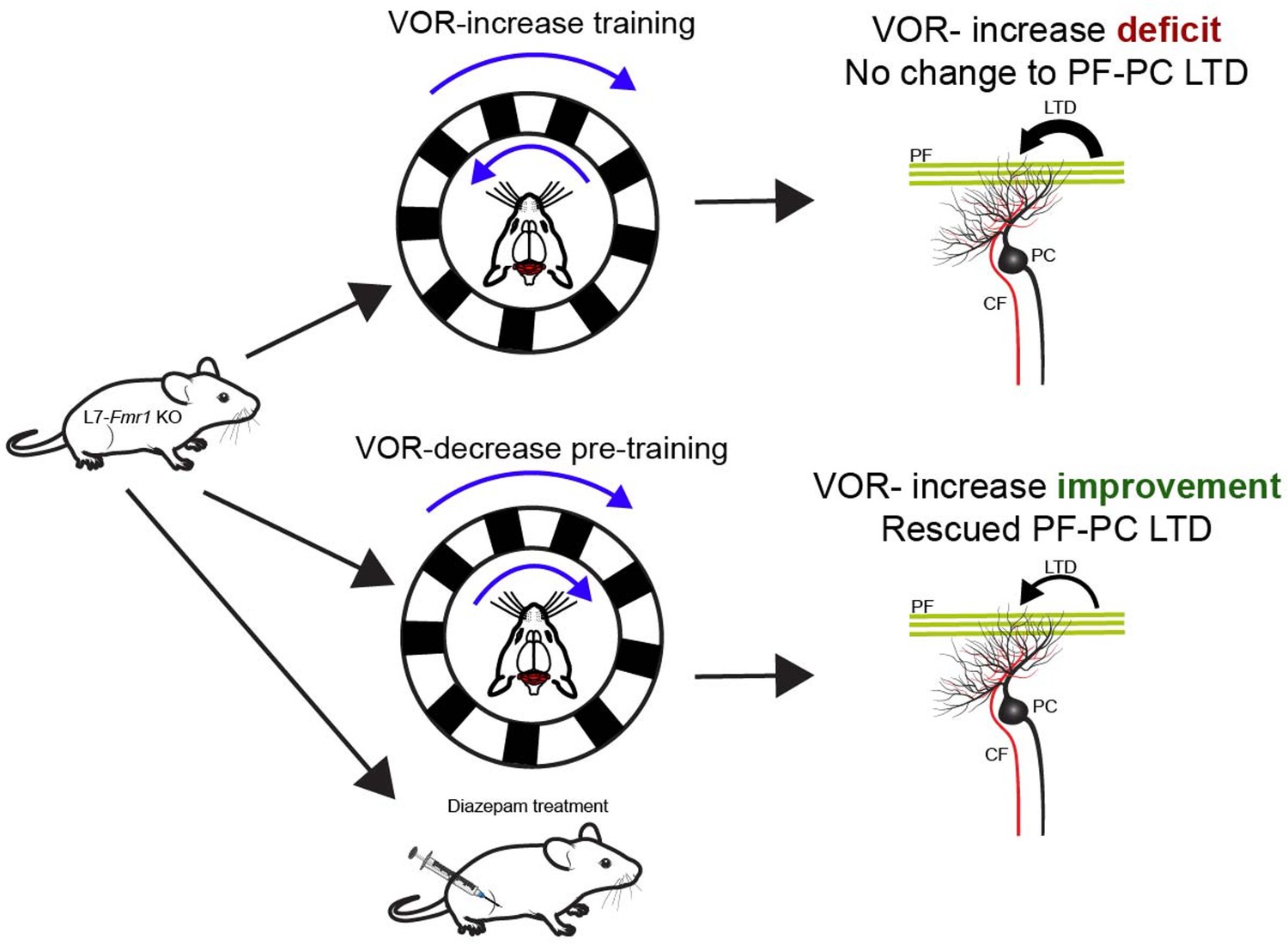
Behavioral and pharmaceutical interventions can rescue vestibulo-ocular reflex increase learning in a mouse model of Fragile X syndrome.
Top: Genetically modified animals (L7-Fmr1 KO mice) whose Purkinje cells carry the mutation that causes Fragile X syndrome show impairments when trained to enhance their vestibulo-ocular reflex (VOR-increase learning). This type of learning relies on the cerebellum (red brain structure). VOR-increase learning involves the animals keeping track of a stimulus (black and white stripes, top blue arrow) rotating in the opposite direction as head movement (bottom blue arrow). The defect is linked to the mice displaying increased long-term depression (LTD; thick black arrow) at the synapses between Purkinje cells (PC; black) and parallel fibers (PF; green). This type of neural plasticity is under the control of both parallel and climbing fibers (CF; red). Bottom: When the VOR-increase learning is preceded by VOR-decrease pre-training (whereby the visual stimulus and head rotate in the same direction) or administration of diazepam, PF-Purkinje cell long-term depression is normalized (thin black arrow) and VOR-increase learning is improved for stimuli rotating at high frequencies.
Image credit: Mouse by Ethan Tyler and Lex Kravitz (CC BY 4.0), mouse head by Luigi Petrucco (CC BY 4.0), Purkinje cells and climbing fibers by Akshay Markanday (CC BY 4.0).
Alternative biological mechanisms may also be involved, however. For instance, the interventions rescued VOR-increase learning when the stimuli used during training were rotating at high frequencies (1Hz), but not at low frequencies (0.5Hz) – possibly because the processing of such frequencies does not rely as much on long-term depression. The mutation associated with Fragile X syndrome may lead to cellular changes or disruptions that were not assessed in this study, and further work should examine whether the structure of Purkinje cells in this mouse model remains unchanged. Follow-up research should also review how these local changes may reshape the long-distance connections between Purkinje cells and neurons outside of the cerebellar microcircuit.
Overall, this work has exciting therapeutic implications for Fragile X syndrome while also expanding our understanding of cerebellar long-term depression. Such knowledge may be crucial as the cerebellum has been implicated in several neurodevelopmental and aging disorders, yet specific treatments that target this structure are still lacking (Wang et al., 2014; Arleo et al., 2024; Iskusnykh et al., 2024).
References
-
Consensus paper: Cerebellum and ageingCerebellum 23:802–832.https://doi.org/10.1007/s12311-023-01577-7
-
A cerebellar learning model of vestibulo-ocular reflex adaptation in wild-type and mutant miceThe Journal of Neuroscience 34:7203–7215.https://doi.org/10.1523/JNEUROSCI.2791-13.2014
-
BookContribution of norepinephrine to cerebellar long-term depression and motor learningIn: Mizusawa H, Kakei S, editors. Cerebellum as a CNS Hub. Springer International Publishing. pp. 337–348.https://doi.org/10.1007/978-3-030-75817-2_16
-
Aging, neurodegenerative disorders, and cerebellumInternational Journal of Molecular Sciences 25:1018.https://doi.org/10.3390/ijms25021018
-
Long-term depressionAnnual Review of Neuroscience 12:85–102.https://doi.org/10.1146/annurev.ne.12.030189.000505
-
Parallel learning and cognitive flexibility impairments between Fmr1 knockout mice and individuals with Fragile X syndromeFrontiers in Behavioral Neuroscience 16:1074682.https://doi.org/10.3389/fnbeh.2022.1074682
Article and author information
Author details
Publication history
Copyright
© 2024, Verpeut
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 455
- views
-
- 44
- downloads
-
- 1
- citation
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 1
- citation for umbrella DOI https://doi.org/10.7554/eLife.100251
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Neural Plasticity: Restoring cerebellar-dependent learning
eLife 13:e100251.
https://doi.org/10.7554/eLife.100251




{kind=link}