Targeted induction of a silent fungal gene cluster encoding the bacteria-specific germination inhibitor fumigermin
- Department of Molecular and Applied Microbiology, Leibniz Institute for Natural Product Research and Infection Biology (HKI), Germany
- Institute of Microbiology, Friedrich Schiller University Jena, Germany
- Department of Biomolecular Chemistry, HKI, Germany
- Leibniz Research Group – Biobricks of Microbial Natural Product Syntheses, HKI, Germany
Abstract
Microorganisms produce numerous secondary metabolites (SMs) with various biological activities. Many of their encoding gene clusters are silent under standard laboratory conditions because for their activation they need the ecological context, such as the presence of other microorganisms. The true ecological function of most SMs remains obscure, but understanding of both the activation of silent gene clusters and the ecological function of the produced compounds is of importance to reveal functional interactions in microbiomes. Here, we report the identification of an as-yet uncharacterized silent gene cluster of the fungus Aspergillus fumigatus, which is activated by the bacterium Streptomyces rapamycinicus during the bacterial-fungal interaction. The resulting natural product is the novel fungal metabolite fumigermin, the biosynthesis of which requires the polyketide synthase FgnA. Fumigermin inhibits germination of spores of the inducing S. rapamycinicus, and thus helps the fungus to defend resources in the shared habitat against a bacterial competitor.
Introduction
In their ecological niche, microorganisms interact with neighboring species in mutualistic or antagonistic ways. For many of their communication processes, they rely on chemical compounds (Mithöfer and Boland, 2016), often belonging to the group of secondary metabolites (SMs) (Keller et al., 2005; Clardy et al., 2009; Brakhage, 2013; Netzker et al., 2015; Keller, 2019). SMs show high chemical diversity, and their encoding genes mostly cluster together in a particular genomic locus (Smith et al., 1990; Trail et al., 1995).
The biosynthesis of SMs is regulated at various levels to ensure production of SMs in a relevant context (Brakhage, 2013). Several environmental signals have been identified as triggers of SM production, such as interactions of neighboring microorganisms (Künzler, 2018). In the artificial setting of a laboratory where natural triggers of SM production are missing, biosynthetic gene clusters are often not expressed and therefore their corresponding SMs remain undetectable (Bergmann et al., 2007; Brakhage, 2013; Rutledge and Challis, 2015). One approach that has proven to be particularly successful in stimulating microbial SM discovery, as well as avoiding redundancy often leading to re-isolation of known compounds, is mimicking the settings of a particular ecological niche through microbial co-culturing (Netzker et al., 2018). In particular, competitive inter-kingdom interactions have been exploited as a trigger for production of bioactive compounds (Watanabe et al., 1982; Cueto et al., 2001; Moree et al., 2013; Khalil et al., 2019). For example, several studies have reported the induction of compound biosynthesis in Aspergillus spp. by Streptomyces (Schroeckh et al., 2009; Wu et al., 2015; Yu et al., 2016) or by Mycobacterium sp. (Jomori et al., 2020). Conversely, downregulation of fungal metabolite biosynthesis by Streptomyces spp. was also reported (Verheecke et al., 2015).
In our previous work, we discovered an inter-kingdom interaction between the soil-dwelling actinomycete Streptomyces rapamycinicus and the filamentous fungus Aspergillus nidulans (Schroeckh et al., 2009). We showed that the silent SM gene cluster encoding biosynthesis of orsellinic acid and its derivatives in A. nidulans is activated on physical contact of the fungus with the bacterium (Schroeckh et al., 2009; Nützmann et al., 2011; Fischer et al., 2018). We subsequently showed that the same bacterial strain also engaged in intimate interplay with the fungus A. fumigatus, in this case triggering expression of the silent polyketide gene cluster encoding the C-prenylated fumicyclines (König et al., 2013). In both instances, the SMs could only be isolated from a fungal-bacterial mixed culture, with no expression of the biosynthetic genes in axenic fungal cultures, which underlines the power of mixed cultures in awakening silenced SM production. In both cases, the ecological function of the produced compounds has remained elusive. However, it is important to unravel general principles of the interactions of microorganisms and to understand functional interactions of microorganisms in microbiomes.
Here, we now report an example where expression of biosynthesis genes of a novel fungal compound is triggered by a bacterium and where the compound then acts on the inducing bacterium.
Results
S. rapamycinicus triggers production of the polyketide fumigermin in A. fumigatus
Based on our previous discoveries that the biosynthesis of novel compounds can be triggered by co-culturing A. nidulans or A. fumigatus with S. rapamycinicus (Schroeckh et al., 2009; König et al., 2013), we analyzed the metabolic profile of the A. fumigatus isolate ATCC 46645 in co-culture with S. rapamycinicus for the presence of further metabolites. Using LC-MS analysis, we detected a new metabolite (1) in the mixed fermentation of A. fumigatus ATCC 46645 and S. rapamycinicus, but not after the addition of supernatant of an S. rapamycinicus culture to the fungal culture (Figure 1A, Figure 1—figure supplement 6). Other streptomycetes such as S. iranensis, S. coelicolor or S. lividans also induced the production of compound 1 during co-cultivation, but to a much lesser extent (Figure 1A). Comparative metabolic analyses by HPLC-HRESI-MS confirmed the presence of high amounts of compound 1 in the extract of the bacterial-fungal co-culture, and trace amounts in that of the axenic fungal culture. For compound 1 a molecular weight of m/z 194 amu was detected. The compound was isolated from the crude extract and its structure was elucidated by NMR analyses. A molecular formula of C11H14O3 was determined by HRESI-MS. The 13C and DEPT135 NMR data (Figure 1—figure supplement 1B, Figure 1—figure supplement 2A) indicated the presence of four methyl and seven double-bonded carbon atoms, six of which proved to be quaternary. The only methine proton (H-7) was shown to be in vicinity to one methyl function (H-8) by the respective H,H-COSY coupling (Figure 1—figure supplement 2B). HMBC couplings of the protons of C-2-CH3 with C-1, C-2 and C-3, and of C-4-CH3 with C-3, C-4 and C-5 revealed the presence of a pyrone ring. HMBC correlations between the C-6-CH3 protons with C-5, C-6 and C-7 and between H-7 and C-5 disclosed the structure and the position of the side chain (Figure 1—figure supplement 3B).
Figure 1 with 6 supplements see all
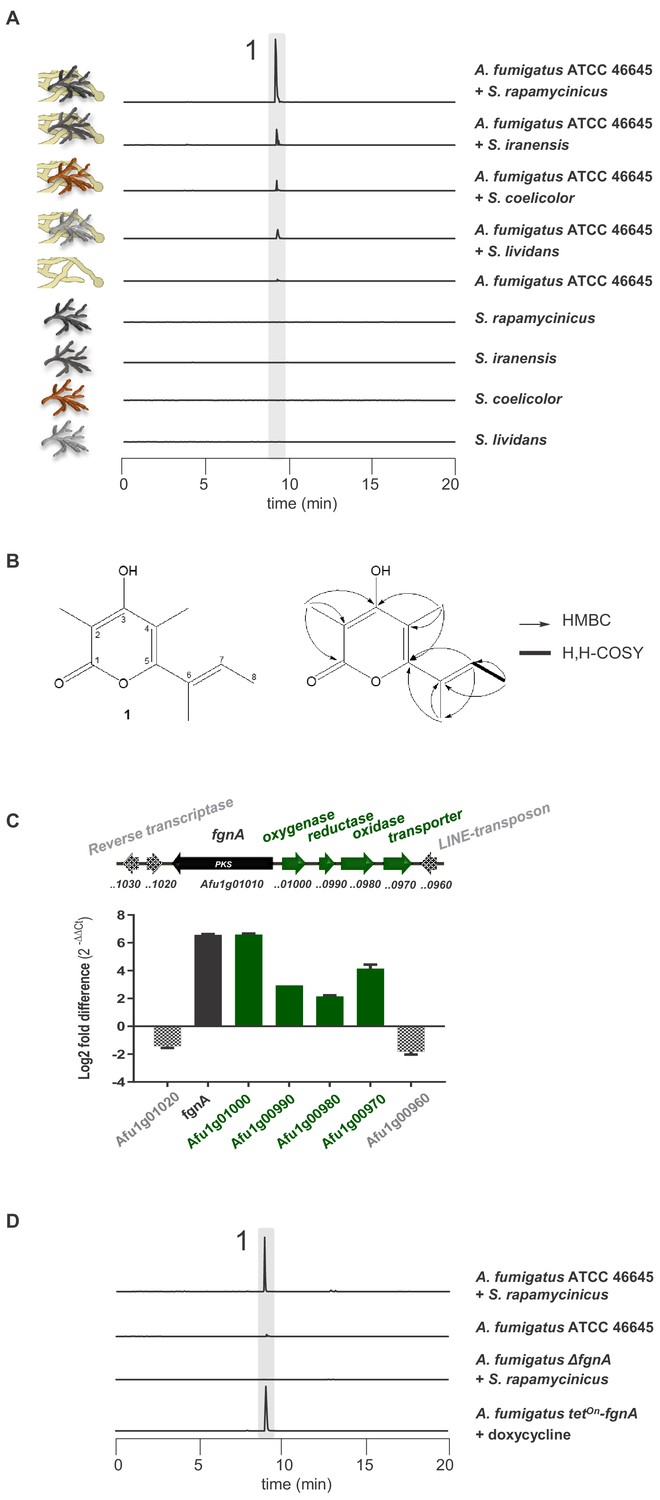
Induction of SM production in A. fumigatus by Streptomyces spp. and identification of the gene responsible for the SM production.
(A) LC-MS analysis of supernatants of A. fumigatus strains after 12 h in axenic or in co-culture with indicated Streptomyces species showing EIC traces for m/z 195 [M+H]+, corresponding to the newly formed compound 1, which is indicated by the highlighted strip. (B) Structure and 2D-NMR correlations of compound 1, fumigermin. (C) Transcription analysis of the A. fumigatus ATCC 46645 fgn cluster and adjacent genes determined by qRT-PCR after 5 h of co-cultivation with S. rapamycinicus. Relative mRNA levels were compared to the β-actin gene transcript levels. The relative expression values are visualized as relative log2 fold difference (2-ΔΔCt) of transcript levels for each gene in co-culture with S. rapamycinicus compared to transcript levels of the respective gene in A. fumigatus ATCC 46645 in axenic culture. Data are representative of three biological replicates with three technical replicates. Error bars indicate standard error of the mean. (D) EIC traces of supernatants of the different A. fumigatus strains in co-cultivation with S. rapamycinicus or under inducing conditions are shown for m/z 195 [M+H]+, which corresponds to fumigermin (1).
-
Figure 1—source data 1
Transcriptional analysis of the fgn cluster.
- https://cdn.elifesciences.org/articles/52541/elife-52541-fig1-data1-v1.xlsx
Compound 1 represents an α-pyrone and was named fumigermin (Figure 1B, Figure 1—figure supplement 4). It is structurally similar to other microbial α-pyrones, including the fungal SMs nectriapyrone from Piricularia oryzae (Motoyama et al., 2019) and gibepyrone produced by Fusarium fujikuroi (Janevska et al., 2016). It is also similar to the bacterial germicidins (Figure 2C) originally isolated from Streptomyces viridochromogens (Aoki et al., 2011), but also produced by Streptomyces species S. coelicolor and S. lividans (Song et al., 2006; Chemler et al., 2012).
Figure 2 with 4 supplements see all

Molecular analysis of the fgnA PKS gene and structure of its biosynthesized product fumigermin.
(A) Heterologous expression of fgnA genes from the indicated A. fumigatus strains in the host A. nidulans. The entire fgn cluster was transferred and expressed in the host A. nidulans. A ‘repaired’ fgnA based on the gene of A1163 was expressed in A. nidulans. (B) PKS gene fgnA in different indicated A. fumigatus strains. SNPs and a gene break are marked. EIC traces of supernatants of the different A. fumigatus strains in co-cultivation with S. rapamycinicus are shown in (A) and (B) for m/z 195 [M+H]+, which corresponds to fumigermin (1). (C) Comparison of the discovered α-pyrone polyketide compound fumigermin (1) and the structurally related bacterial pyrones germicidin A (2), germicidin B (3), nectriapyrone (4) and gibepyrone (5). (D) FgnA enzyme domains and proposed biosynthesis of fumigermin.
Fumigermin biosynthesis requires the fgnA polyketide synthase gene
The induced production of a fungal metabolite prompted us to monitor the expression of potential biosynthesis genes in co-culture. The elemental composition of the metabolite revealed by its HRESI-MS-deduced molecular formula (195.1015 [M+H]+, calcd. C11H15O3 195.1016) showed a lack of nitrogen atoms and therefore pointed towards a possible polyketide origin of the compound. Additionally, the structure of the compound suggested that its biosynthesis is catalyzed by a partially reducing polyketide synthase (PKS). Bioinformatic inspection of the genome (Bignell et al., 2016) indicated that only a few clusters matched this prediction. One of these clusters, whose products are so far unknown, was the uncharacterized gene cluster Afu1g00970-Afu1g01010 (Supplementary file 1), containing the putative PKS gene Afu1g01010.
To verify whether the selected genes are involved in biosynthesis of the detected metabolite produced in the bacterial-fungal interaction, we analyzed expression of cluster genes Afu1g00970-Afu1g01010 of A. fumigatus and their 3’ and 5’ adjacent genes by qRT-PCR after 5 h of co-culture. As indicated in Figure 1C, the steady-state mRNA levels of the putative cluster genes of A. fumigatus ATCC 46645, including gene Afu1g01010, were markedly upregulated during co-cultivation of A. fumigatus ATCC 46645 with S. rapamycinicus. To unequivocally prove that the identified PKS gene Afu1g01010 encodes the key enzyme involved in the biosynthesis of fumigermin, we deleted Afu1g01010 in the genome of A. fumigatus ATCC 46645 (Figure 1—figure supplement 5). LC-MS monitoring of the co-cultivation of the resulting deletion mutant with S. rapamycinicus demonstrated that production of fumigermin (1) was fully abolished (Figure 1D), confirming that the PKS gene Afu1g01010 is involved in its biosynthesis. This was further proven by overexpressing Afu1g01010, hereby named fgnA, using the inducible tetOn promoter system in A. fumigatus ATCC 46645, which led to the appearance of fumigermin under inducing conditions in the absence of bacteria (Figure 1D). Taken together, these data clearly prove that the identified fgnA gene is strictly required for biosynthesis of the novel metabolite fumigermin (1).
The fgnA polyketide synthase gene is essential for production of fumigermin
To characterize the biosynthetic pathway of fumigermin, we expressed the fgnA gene from strain ATCC 46645 heterologously in A. nidulans, under control of the constitutive gpdA promoter. Metabolic profiling of the culture of A. nidulans overexpressing the fgnA gene of strain ATCC 46645 revealed the formation of 1, a metabolite identical to fumigermin which had been previously detected during the co-cultivation of A. fumigatus and S. rapamycinicus (Figure 2A). This finding suggested that fumigermin, that is compound 1, is a direct product of the encoded polyketide synthase FgnA of A. fumigatus.
To prove that the encoded putative tailoring enzymes are non-essential for biosynthesis of fumigermin, we transferred the entire fgn cluster from A. fumigatus ATCC 46645 to A. nidulans RMS011. For this purpose, we applied our previously developed heterologous expression system allowing for expression of several genes from a polycistronic mRNA in a eukaryotic host (Unkles et al., 2014; Hoefgen et al., 2018). This expression system is based on post-translational cleavage of proteins at the viral 2A peptide sequences (Whitton et al., 2005). Briefly, all cluster genes were cloned in a plasmid such that they were all separated by 2A peptide-encoding sequences. To facilitate selection of transformants harboring such plasmids, we employed a gene encoding a split-Venus fluorescent protein that, in addition, contained a nuclear localization sequence. In the plasmid, the gene cluster was flanked by two genes, each encoding a half of the Venus gene. Consequently, when all genes of the plasmid are expressed, the two Venus halves (N+C) are also produced and bind to each other to form a functional Venus fluorescent protein. As a result, fluorescence in the fungal nucleus indicates correct translation of the encoded proteins. The entire cluster construct was placed under the control of the tetOn promoter, allowing inducible expression of all genes from a polycistronic mRNA (Figure 2—figure supplement 1A). Although all cluster proteins were produced in the heterologous strain, as evidenced by the fluorescent signal localized to the nuclei (Figure 2—figure supplement 1B) and additionally supported by label-free proteomic analysis (Figure 2—figure supplement 1C), no further compound, other than fumigermin, was detected in the heterologous expression strain (Figure 2A, Figure 2—figure supplement 1B). This result indicates that the additional enzymes do not modify the chemical product of FgnA, even when co-expressed with the functional FgnA PKS.
The partially reducing FgnA synthesizes fumigermin via a PKS I-derived pathway
The deduced amino acid sequence of FgnA analyzed by CD-search (Marchler-Bauer and Bryant, 2004) revealed the presence of a ketosynthase (KS), acyltransferase (AT), dehydratase (DH), methyl transferase (MT), ketoreductase (KR) and an acyl-carrier protein (ACP) domain, classifying this enzyme as a partially reducing PKS (Cox, 2007). Isotope labeling studies using (2-13C)acetate and (methyl-13C)methionine showed that eight carbon atoms of fumigermin originate from acetate, while the remaining three carbons derived from S-adenosyl methionine, agreeing with the existence of an MT domain in the PKS (Figure 1—figure supplement 4). The conserved domains together with the precursor studies led to the proposed model for fumigermin biosynthesis by FgnA (Figure 2D).
In A. fumigatus a functional FgnA is restricted to strain ATCC 46645
Among Aspergillus spp., the fgn cluster is unique to A. fumigatus and the closely related species A. novofumigatus (Figure 2B). Moreover, the cluster is rarely found in the different sequenced A. fumigatus genomes, which is consistent with a previously reported analysis of clusters in various A. fumigatus isolates (Lind et al., 2017). Amongst the A. fumigatus strains harboring the fgn cluster, production of fumigermin was restricted to strain ATCC 46645. We could not detect fumigermin in a co-culture of S. rapamycinicus with other A. fumigatus strains, such as Af293 and A1163, or A. novofumigatus IBT 16806 (Figure 2B). To determine the reason for the missing compound production in strains Af293, A1163 and IBT 16806, we analyzed their respective genome sequences retrieved from the NCBI database, with a focus on the fgnA gene. For strain A1163, the available genome sequence did not cover the area around the putative Afu1g01010 cluster. Thus, we amplified the fgnA gene of A1163 by PCR and sequenced it. As indicated by the gene sequences, the corresponding fgnA gene of strains Af293 and IBT 16806 contained several missense SNPs, while that of strain A1163 had undergone a chromosomal breakage around 800 bp from the start codon of fgnA, and contained a C → T missense mutation at position 272 (Figure 2B, Figure 2—figure supplement 3). Thus, these genes appeared to be non-functional.
To further verify the assumption that the fgnA genes are not functional in several A. fumigatus strains, we overexpressed the fgnA genes of both the Af293 and A1163 strains in A. nidulans. As predicted, neither FgnA PKS was able to produce fumigermin (Figure 2A). To test whether the repair of the SNP in a A1163-derived PKS gene would rescue the function of the biosynthetic enzyme, the two PCR-derived DNA fragments covering the A1163 fgnA gene were first ligated in a plasmid and then the missense SNP was corrected. The heterologous overexpression of the resulting ‘repaired’ fgnA gene in A. nidulans under the control of the gpdA promoter led to formation of the expected polyketide metabolite fumigermin, confirming that this reconstruction functionally rescued the ‘inactive’ A1163 fgnA gene and led to production of the previously observed metabolite derived from the fgnA gene (Figure 2A).
The fumigermin synthase gene is spread among distantly related fungal classes
To identify potential orthologs of FgnA and to understand its relation to other fungal PKS, we performed Uniprot database searches and phylogenetic analyses. The obtained data indicate that additional orthologs of the entire fgn cluster are found in distantly related fungi such as the major pathogen of wheat Parastagonopora nodorum from the fungal taxon Dothideomycetes (Hane et al., 2007), the bat pathogen Pseudogymnoascus pannorum which belongs to the Leotiomycetes (Minnis and Lindner, 2013) and fungi of the Sodariomycetes such as the parasitic microfungus Escovopsis weberi (Reynolds and Currie, 2004) and the endophyte Hypoxylon sp. (Hsieh et al., 2005; Supplementary file 5). As these microorganisms are derived from different fungal classes and occupy diverse habitats, it is conceivable that the fgn cluster was spread between different fungal species through horizontal gene transfer.
Surprisingly, a similarity search based on the protein sequence of FgnA did not retrieve the stand-alone PKS enzymes Nec1 and Gpy1, which catalyze the biosynthesis of the structurally similar microbial SMs nectriapyrone (Figure 2—figure supplement 4; Janevska et al., 2016) and gibepyrone (Motoyama et al., 2019), respectively. Despite also being classified as fungal iterative type I PKS, Nec1 and Gpy1 are not orthologs of FgnA, and show low similarity to FgnA at the amino acid level. Moreover, Nec1 and Gpy1 also constitute interesting examples of stand-alone PKS enzymes that produce compounds similar to fumigermin and that do not require additional cluster-encoded enzymes.
Fumigermin reversibly inhibits the germination of spores of the inducing S. rapamycinicus
Fumigermin displayed no remarkable antimicrobial effects, except for a modest inhibitory activity against MRSA (data not shown). To learn more about the potential ecological role of fumigermin, we investigated its effects on the inducing streptomycetes. Fumigermin is structurally related to germicidins, which have been shown to function as auto-regulators of spore germination, that is they inhibit the germination of the producing streptomycetes (Aoki et al., 2011). We therefore tested whether fumigermin has a similar activity, which would provide an explanation for the interaction between A. fumigatus and the streptomycetes. As shown in Figure 3A and B, the number of germinated S. rapamycinicus spores was drastically reduced by fumigermin in a dose-dependent manner. A comparison of fumigermin and germicidin B demonstrated that both compounds showed the same anti-germination effect against spores of S. rapamycinicus (Figure 3C). Moreover, fumigermin also inhibited germination of S. rapamyinicus spores out of solid medium (Figure 3D). To reveal whether the inhibition of spore germination was reversible, we incubated S. rapamycinicus spores with fumigermin for 90 h, and subsequently washed and re-cultivated the spores in fresh media. As shown in Figure 3E and Figure 3—figure supplement 1, the spores' ability to germinate and form a mycelial network was regained. Thus, fumigermin is a reversible germination inhibitor, which keeps S. rapamycinicus spores in a dormant state and leaves them ungerminated as long as the compound is present. Notably, fumigermin could be re-purified from 90 h-old supernatant of a fumigermin-treated S. rapamycinicus spore sample, and further re-used to inhibit germination of fresh S. rapamycinicus spores (Figure 3—figure supplement 1). This finding points towards high stability of the compound.
Figure 3 with 3 supplements see all

Activity of fumigermin against germination of S. rapamycinicus spores.
(A) 107 spores of S. rapamycinicus were inoculated in 500 µL M79 medium in a 48-well plate and treated with increasing fumigermin concentrations. Image taken after 5 days of growth at 28°C, 500 rpm. Magnified areas zoom in on an untreated sample, sample treated with 50 µg/mL or 175 µg/mL fumigermin and a control sample containing medium with no spores. (B) OD595 measurements were taken for each well using a Tecan device. The mean OD595 value of the untreated S. rapamycinicus samples was set as 100% germination, and used to calculate the relative germination percentage of the fumigermin-treated samples. (C) Graph of 90-h-long monitoring of S. rapamycinicus spore germination inhibition at 28°C using germicidin B and fumigermin (250 μg/mL). Controls were treated with MeOH or remained untreated. An increase in the OD595 value is indicative of S. rapamycinicus spore germination. Data in (B, C) correspond to three biological samples, each with three technical replicates. Error bars indicate standard error of the mean. (D) Qualitative inhibition of formation of S. rapamycinicus aerial mycelia by fumigermin. Oatmeal agar plates with embedded S. rapamycinicus spores were point-inoculated with different concentrations of fumigermin: (1) 12.5 μg, (2) 25 μg, (3) 50 μg, (4) 75 μg, (5) 100 μg, (6) 250 μg and incubated for 1 week at 28°C. Left, zoom of area (1). (E) Reversibility of germination inhibition. S. rapamycinicus spores were incubated with 250 μg/mL fumigermin. After 5 days, the spores did not germinate; when recovered, washed and placed in fresh medium, the spores grew and formed mycelia.
-
Figure 3—source data 1
OD595 measurement of S. rapamycinicus growth.
- https://cdn.elifesciences.org/articles/52541/elife-52541-fig3-data1-v1.xlsx
Because S. lividans and S. coelicolor only triggered production of moderate fumigermin titers, we tested whether fumigermin also affected the growth of these streptomycetes. The germination of S. coelicolor and S. lividans was only slightly inhibited when treated with fumigermin. Moderate germination inhibition was observed for S. coelicolor treated with the rather high concentration of 1000 µg/mL fumigermin (Figure 3—figure supplement 2). The inhibitory effect was lost after 3 days of incubation with the compound, suggesting that fumigermin might be degraded by this Streptomyces spp. To test this assumption, the purified metabolite was supplemented to growing cultures of S. rapamyinicus, S. iranensis, S. lividans and S. coelicolor, and a control culture of Pseudomonas aeruginosa PAO1. After 24 h of incubation, the levels of fumigermin were determined by LC-MS (Figure 3—figure supplement 3). The analysis showed that none of the streptomycetes tested was able to degrade fumigermin, which was in contrast to the P. aeruginosa PAO1 control culture where fumigermin was almost depleted in the culture supernatant. As expected, fumigermin had no obvious effect on the fitness of the producing isolate A. fumigatus ATCC 46645 at least when cultivated in monoculture (data not shown).
Discussion
Fumigermin reversibly inhibits germination of susceptible Streptomyces strains
Employing interactions of microorganisms to activate cryptic SM gene clusters has been successfully established as a methodology to identify novel compounds (Netzker et al., 2018). Here, we uncovered a novel fungal SM, production of which was induced during the interaction of A. fumigatus with S. rapamycinicus. It is reasonable to assume that SMs are produced for the benefit of the producer microorganism in a specific niche (Macheleidt et al., 2016). However, the ecological benefit for the producer has only been established for a few cases (Notz et al., 2002; Tanaka et al., 2005; Khalil et al., 2019). For instance, A. flavus produces aflavinines as protection against the beetle Carpophilus hemipterus (Gloer and Truckenbrod, 1988). The fungal endophyte Epichloë festucae produces peramine to protect its plant host from feeding by Listronotus bonariensis (Tanaka et al., 2005). Moreover, fusaric acid produced by Fusarium graminearum represses production of an antifungal compound in Pseudomonas fluorescens CHA0 (Notz et al., 2002). Consistently, fumigermin is not produced in an axenic A. fumigatus ATCC 46645 culture, but is produced in high amounts and excreted by A. fumigatus ATCC 46645 when co-cultured with S. rapamycinicus.
Aspergillus spp., in particular A. fumigatus, and Streptomyces spp. have been isolated previously from a soil sample and found to share the same habitat (Alani et al., 2012; Khalil et al., 2019), making it likely that this laboratory scenario is also relevant for the ecological niche of the analyzed microorganisms. However, there is little insight into the ecology of bacterial-fungal interactions yet. In the case of A. fumigatus ATCC 46645 and S. rapamycinicus reported here, the two microorganisms are apparently interacting in an antagonistic manner, because the fungus is triggered by the bacterium to produce the germination inhibitor fumigermin to protect its habitat’s resources (Figure 4). As fumigermin’s inhibitory activity is reversible, it is likely that the fungus does not intend to kill the bacterium.
Figure 4

Model of the interaction between S. rapamycinicus and A. fumigatus ATCC 46645, which results in production of fumigermin.
During the co-cultivation of the two microorganisms, the bacterium S. rapamycinicus induces expression of the fgn gene cluster of the fungus A. fumigatus ATCC 46645, which leads to production of the SM fumigermin. Fumigermin is secreted and further inhibits germination of S. rapamycinicus spores.
In this work, several Streptomyces spp. were found to induce fumigermin biosynthesis. Whereas S. iranensis is phylogenetically closely related to the fumigermin-inducer S. rapamycinicus, S. coelicolor and S. lividans are rather distantly related to S. rapamycinicus (Nouioui et al., 2018). This phylogenetic relationship mirrors the extent of induction of fumigermin production in A. fumigatus and also correlates with the activity of fumigermin against the inducing Streptomyces spp.. More specifically, fumigermin was highly active against the main fumigermin inducers S. rapamycinicus and its close relative S. iranensis (data not shown). On the other hand, S. coelicolor and S. lividans triggered production of lower levels of fumigermin and were not susceptible to fumigermin. Our data on the degree of induction and the susceptibility of Streptomyces species thus indicate the specificity of the interaction of S. rapamycinicus/S. iranensis and A. fumigatus. S. coelicolor and S. lividans are known producers of the structurally related α-pyrone metabolites germicidins (Song et al., 2006; Chemler et al., 2012). It is thus likely that these streptomycetes harbor an intrinsic resistance mechanism against germination-inhibiting α-pyrone compounds. Because none of the Streptomyces species analyzed here degraded fumigermin, it is possible that other resistance mechanisms such as specific export of germicidins, are responsible for the intrinsic resistance.
Germicidins are autoregulatory inhibitors of germination of dormant spores in a Streptomyces population. Streptomyces germ tubes secrete germicidins, which presumably block the activity of a membrane located Ca2+-dependent ATPase (Eaton and Ensign, 1980; Grund and Ensign, 1985; Aoki et al., 2011). The role of germicidins for the ecology of streptomycetes is a matter of debate (Čihák et al., 2017). It is conceivable that A. fumigatus has acquired the ability to biosynthesize a functionally similar compound and is exploiting this effect to its advantage during interaction with the bacteria, as means of protection of the habitat’s resources from competitor microorganisms.
The biosynthesis routes of α-pyrone-based germination inhibitors possibly developed by convergent evolution
The structural similarity between fumigermin and germicidins and their same biological activity initially suggested that the compounds share a common biosynthetic origin. However, germicidins and fumigermin are products of pathways that are different. Whereas germicidins are produced by a type II fatty acid synthase coupled with a bacterial type III PKS that uses precursors derived from fatty acid biosynthesis (Figure 5 and Chemler et al., 2012), fumigermin biosynthesis involves a fungal iterative type I PKS. This finding makes a horizontal gene transfer from bacteria to fungi unlikely and rather points to a possible case of convergent evolution of the same lead structure in the bacterial and fungal kingdom.
Figure 5
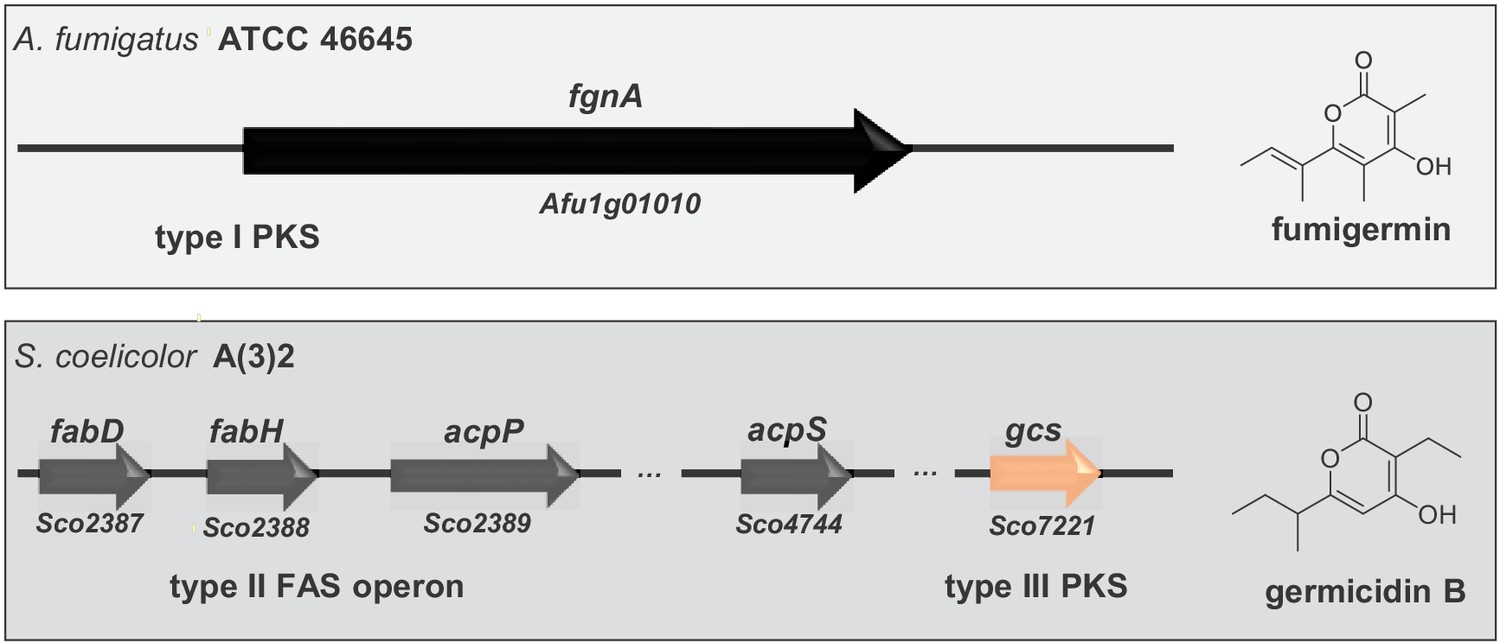
Alignment of genes involved in biosynthesis of the fungal compound fumigermin and the bacterial compound germicidin B.
Germicidin synthase is a type III bacterial PKS, while FgnA is a fungal iterative type I PKS. The synthesis of germicidins is coupled with fatty acid biosynthesis in S. coelicolor - the branched precursors are synthesized by FabD and FabH, after which they are further loaded onto and modified by Gcs (Chemler et al., 2012). In case of A. fumigatus, FgnA catalyzes the condensation of malonyl-CoA units to an acetyl-CoA starter unit independently of the fatty acid biosynthesis pathway. There is no striking similarity between Gcs and FgnA.
A. fumigatus-specific fumigermin PKS gene is sufficient for germicidin production despite being located in a cluster
It is remarkable that the five cluster genes including fgnA are conserved in distantly related fungi, supporting the assumption of a fgn cluster. It is thus possible that originally all five genes were functional, but that the genes encoding tailoring enzymes became inactive from mutations, similar to the case of the fgnA gene in strains A1163 and Af293. In A. fumigatus, the fgn locus has been shown to be particularly prone to rearrangements and breakages, possibly as a result of being flanked by potential transposons (Figure 1C, Supplementary file 1). Consistently, Lind et al. have shown that the cluster ‘jumped’ to different genomic locations in three of the six analyzed cluster-containing strains, in one case being located inside the fumagillin/pseurotin supercluster (Wiemann et al., 2013; Lind et al., 2017). Additionally, loss of gene clusters apparently tends to occur at higher rates when they are located near telomere ends (Young et al., 2015), which might explain that the fgn cluster is only present in a few A. fumigatus isolates.
Collectively, a rational explanation regarding the fate of the fgn cluster is that the modifications suffered as a consequence of its chromosomal location, or because of repeated transpositions driven by the putative cluster-flanking mobile elements, have led to breakages and SNP formation and a consequent loss of cluster functionality in most of the A. fumigatus strains. Our data so far indicate that the isolate ATCC 46645 is the only Aspergillus sp. in which the cluster was maintained, even though the tailoring enzymes are apparently non-functional (Figure 2—figure supplement 1). The production of fumigermin likely supported the conservation of FgnA, as it was advantageous for the particular isolate in its interaction with streptomycetes (Figure 4).
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Aspergillus fumigatus) | Af293 | Nierman et al., 2005 | Wild-type strain, MAT1-2 | |
| Strain, strain background (Aspergillus fumigatus) | A1163 | da Silva Ferreira et al., 2006 | Wild-type strain, MAT1-1 | |
| Strain, strain background (Aspergillus fumigatus) | ATCC 46645 | Langfelder et al., 1998 | Wild-type strain, MAT1-1 | |
| Strain, strain background (A. novofumigatus) | IBT 16806 | Hong et al., 2005 | Wild-type strain | |
| Strain, strain background (Aspergillus fumigatus) | ΔfgnA | This paper | ATCC 46645 background, ptrA::fgnA, PTR | |
| Strain, strain background (Aspergillus fumigatus) | tetOn-fgnA | This paper | ATCC 46645 background, tetOn-fgnA, PTR | |
| Strain, strain background (Aspergillus nidulans) | RMS011 | Stringer et al., 1991 | pabaA1, yA2; ΔargB::trpCΔB, trpC801, veA1 | |
| Strain, strain background (Aspergillus nidulans) | fgnA_ATCC | This paper | RMS011 background, pUC18-gpdA- fgnA_ATCC, ArgB2+ | |
| Strain, strain background (Aspergillus nidulans) | fgnA_Af293 | This paper | RMS011 background, pUC18-gpdA- fgnA_Af293, ArgB2+ | |
| Strain, strain background (Aspergillus nidulans) | fgnA_A1163_repaired | This paper | RMS011 background, pUC18-gpdA-fgnA_A1163*, ArgB2+ | |
| Strain, strain background (Aspergillus nidulans) | fgn_cluster | This paper | RMS011 background, fgnA,Afu1g01000,Afu1g00990,Afu1g00980,Afu1g00970, ArgB2+ | |
| Strain, strain background (Streptomyces rapamycinicus) | ATCC 29253 | Kumar and Goodfellow, 2008 | Wild-type strain | |
| Strain, strain background (Streptomyces iranensis) | HM35 | Hamedi et al., 2010 | Wild-type strain | |
| Strain, strain background (Streptomyces lividans) | PM02 | van Dissel et al., 2015 | Wild-type strain | |
| Strain, strain background (Streptomyces coelicolor) | A(3)2 | Erikson, 1955 | Wild-type strain | |
| Commercial assay or kit | NEB HiFi | New England Biolabs, Frankfurt, Germany | E2621L | |
| Commercial assay or kit | Universal RNA Purification Kit | Roboklon, Berlin, Germany | E3598 | |
| Chemical compound, drug | germicidin B | Cayman Chemical, Ann Arbor, USA | CAS number 150973-78-7 | |
| Chemical compound, drug | digoxigenin-11- dUT | Jena BioScience, Jena, Germany | Catalog number NU-803-DIGXS | |
| Software, algorithm | GraphPad Prism 5 | GraphPad Software Inc,La Jolla, USA | RRID:SCR_002798 | |
| Software, algorithm | Thermo Xcalibur Qual Browser | Thermo Scientific, Dreieich, Germany | ||
| Software, algorithm | Proteome Discoverer (PD) 2.2 | Thermo Scientific, Dreieich, Germany | RRID:SCR_014477 | |
| Software, algorithm | Shimadzu Class-VP software (version 6.14 SP1) | Shimadzu, Duisburg, Germany | 6.14 SP1 | |
| Other | Hoechst 34580 | Thermo Fischer Scientific, Dreieich, Germany | H21486 |
Microorganisms, media and cultivation
Request a detailed protocolMicroorganisms used in this study are listed in the Key Resources Table. A. fumigatus and A. nidulans strains were cultivated in Aspergillus minimal medium (AMM) at 37°C, 200 rpm (Brakhage and Van den Brulle, 1995). Fungal pre-cultures were inoculated with 4 × 108 spores per mL. For A. fumigatus tetOn-fgnA and A. nidulans fgn_cluster, 10 µg/mL doxycycline was used to induce the tetOn inducible system. For A. nidulans strains, the culture medium was supplemented with arginine (871 µg/mL) and p-aminobenzoic acid (3 mg/L) when needed. S. rapamycinicus and S. coelicolor were cultivated on oatmeal agar plates (Shirling and Gottlieb, 1966) for sporulation and in M79 medium (Prauser and Falta, 1968) for liquid cultivation. S. lividans was cultivated on mannitol soya flour (MS) agar plates (Hobbs et al., 1989) to obtain spores. For cultivation in liquid media tryptone soy broth (TSB) was employed (Kieser et al., 2000) with 10% (w/v) sucrose. Cultures were propagated at 28°C and shaken at 160 rpm. For co-cultivation experiments, mycelia of A. fumigatus (~16 h old) were separated from the medium using Miracloth (Merck Millipore, Darmstadt, Germany), placed in fresh AMM and inoculated with 1/20 vol of the streptomycete culture as previously described (Schroeckh et al., 2009), or with 1/20 vol of the streptomycete culture supernatant, filtered through a 0.22 µm filter. RNA extraction for expression analysis was performed after 5 h of co-cultivation, while samples for LC-MS analysis were taken after 12 h of co-cultivation.
Extraction of fungal compounds and LC-MS analysis
Request a detailed protocolThe culture broth containing fungal mycelium with and without bacteria was homogenized using an ULTRA-TURRAX (IKA-Werke, Staufen, Germany). Homogenized cultures were extracted twice with a total of 100 mL ethyl acetate, dried with sodium sulfate and concentrated under reduced pressure. For LC-MS analysis, the dried extracts were dissolved in 1 mL of methanol and loaded onto an ultrahigh-performance liquid chromatography (LC)–MS system consisting of an UltiMate 3000 binary rapid-separation liquid chromatograph with photodiode array detector (Thermo Fisher Scientific, Dreieich, Germany) and an LTQ XL linear ion trap mass spectrometer (Thermo Fisher Scientific, Dreieich, Germany) equipped with an electrospray ion source. The extracts (injection volume, 10 μl) were analyzed on a 150 mm by 4.6 mm Accucore reversed-phase (RP)-MS column with a particle size of 2.6 μm (Thermo Fisher Scientific, Dreieich, Germany) at a flow rate of 1 mL/min, with the following gradient over 21 min: initial 0.1% (v/v) HCOOH-MeCN/0.1% (v/v) HCOOH-H2O 0/100, which was increased to 80/20 in 15 min and then to 100/0 in 2 min, held at 100/0 for 2 min, and reversed to 0/100 in 2 min.
Compound isolation and structure elucidation by NMR
Request a detailed protocolCompounds were isolated by preparative HPLC using a Shimadzu LC-8a series HPLC system with DAD and a Synergi-Fusion RP18 column (Phenomenex, Aschaffenburg, Germany) at a flow rate of 10 mL/min and a gradient system of 0.01% (v/v) TFA (H2O) and MeCN starting at 10% (v/v) MeCN leading to 90% in 20 min. HPLC-HRESI-MS measurements were performed using a Q Exactive hybrid quadrupole-orbitrap mass spectrometer with an electrospray ion source (capillary voltage 3.2 kV, capillary temperature 250°C) coupled to an Accela HPLC system (Thermo Fisher Scientific, Bremen, Germany). HPLC conditions: C18 column (Accucore C18 2.6 µm 100 mm x 2.1 mm) and gradient elution (acetonitrile/H2O (each containing 0.1% (v/v) formic acid) 5/95 going up to 98/2 in 10 min, then 98/2 for another 4 min; flow rate 0.2 mL/ min). NMR spectra were recorded on a Bruker Avance DRX 600 instrument. Spectra were normalized to the residual solvent signals.
Analysis of heterologously expressed A. fumigatus fgn cluster proteins
Request a detailed protocolProtein preparation and LC-MS/MS analysis and database search for identification of proteins were essentially performed as previously described (Baldin et al., 2015), except the following changes: LC gradient elution was as follows: 0 min at 4% B, 5 min at 5% B, 30 min at 8% B, 60 min at 12% B, 100 min at 20% B, 120 min at 25% B, 140 min at 35% B, 150 min at 45% B, 160 min at 60 %B, 170–175 min at 96% B, 175.1–200 min at 4% B. Mass spectrometry analysis was performed on a QExactive HF instrument (Thermo Scientific, Dreieich, Germany) at a resolution of 120,000 FWHM for MS1 scans and 15,000 FWHM for MS2 scans. Tandem mass spectra were searched against the FungiDB database (2020/01/07 https://fungidb.org/common/downloads/Current_Release/AnidulansFGSCA4/fasta/data/FungiDB-46_AnidulansFGSCA4_AnnotatedProteins.fasta) of Aspergillus nidulans (FGSC A4) and the protein sequences Afu1g01010, Afu1g01000, Afu1g00990, Afu1g00980, and Afu1g00970 of Aspergillus fumigatus Af293 using Proteome Discoverer (PD) 2.2 (Thermo Scientific, Dreieich, Germany) and the algorithms of Sequest HT (version of PD2.2), Mascot 2.4, and MS Amanda 2.0. Modifications were defined as dynamic Met oxidation and protein N-term acetylation as well as static Cys carbamidomethylation.
Preparation of chromosomal DNA and RNA purification
Request a detailed protocolA. fumigatus and A. nidulans genomic DNA was isolated as previously described (Schroeckh et al., 2009). A. fumigatus total RNA was purified with the Universal RNA Purification Kit (Roboklon, Berlin, Germany) according to the manufacturer’s instructions.
Quantitative RT-PCR (qRT-PCR)
Request a detailed protocolReverse transcription of 5 µg RNA was performed with RevertAid Reverse Transcriptase (Thermo Fisher Scientific, Darmstadt, Germany) for 3 h at 46°C. qRT-PCR was performed as described before (Schroeckh et al., 2009). The A. fumigatus β-actin gene (Afu6g07470) served as an internal standard for calculation of expression levels as previously described (Schroeckh et al., 2009). Primers used for the amplification of probes are listed in Supplementary file 3.
Generation of an A. fumigatus fgnA deletion strain
Request a detailed protocolThe transformation cassettes for the A. fumigatus fgnA deletion strain were constructed essentially as previously described (Szewczyk et al., 2006). 2000 bp sequences homologous to the regions upstream and downstream of fgnA (Afu1g01010) were amplified using primer pairs P1/P2 and P3/P4. The pyrithiamine (ptrA) resistance cassette was amplified from plasmid pSK275 (Szewczyk and Krappmann, 2010) with the primer pair P5/P6. The above PCR-amplified DNA fragments were assembled together with the HindIII-linearized pUC18 vector using the NEBuilder HiFi DNA Assembly Master Mix (New England Biolabs, Frankfurt, Germany) according to the manufacturer’s instructions, yielding plasmid pUC18-ptrA::fgnA. The final fgnA deletion construct was generated by PCR amplification of the 2 kb upstream and downstream coding regions of fgnA flanking the pyrithiamine resistance cassette from plasmid pUC18-ptrA::fgnA using primers P1/P4. A. fumigatus strain ATCC 46645 was transformed with the resulting linear DNA product as previously described (Unkles et al., 2014). Colonies of transformant strains were selected on AMM agar plates supplemented with 0.1 µg/mL pyrithiamine (Sigma-Aldrich, Hamburg, Germany).
Generation of an inducible A. fumigatus fgnA overexpressing strain
Request a detailed protocolFor overexpression of fgnA, the tetracycline-controlled transcriptional activation system (tetOn) was used (Helmschrott et al., 2013) and integrated in locus directly upstream of the fgnA gene. For this purpose, 2000-bp flanking regions were amplified from A. fumigatus ATCC 46645 genomic DNA with the primer pairs P7/P8 and P9/P10, while the tetOn-system was amplified from plasmid pSK562 (Sven Krappmann, personal communication) with primers P11/P12 and the ptrA cassette was amplified from plasmid pSK275 with the primer pair P5/P6. The above generated PCR fragments were assembled together with the HindIII-linearized pUC18 vector using the NEBuilder HiFi DNA Assembly Master Mix (New England Biolabs, Frankfurt, Germany) according to the manufacturer’s instructions, yielding plasmid pUC18-tetOn-fgnA. The final transformation construct was generated by PCR amplification from plasmid pUC18-tetOn-fgnA using primers P7/P10. The obtained linear DNA fragment, comprising the 2 kb coding regions upstream and downstream of the fgnA (Afu1g1g01010) ATG start codon, flanking the tetOn promoter and the ptrA cassette, was used for transformation of A. fumigatus ATCC 46645. Transformants were selected on AMM agar plates supplemented with 0.1 µg/mL pyrithiamine (Sigma-Aldrich, Hamburg, Germany).
Heterologous expression of genes of the fgn cluster in A. nidulans
Request a detailed protocolFor overexpression of fgnA in A. nidulans, the fgnA gene was PCR-amplified with primers P13/P14 from either A. fumigatus Af293 or ATCC 46645 and fused with the constitutively active gpdA promoter of A. nidulans amplified together with the pUC18 backbone from plasmid pUC18-tetOn-basR (Fischer et al., 2018) with primer pair P15/P16. The A. nidulans argB2 gene was used as a selectable marker to complement the arginine auxotrophy of the A. nidulans RMS011 wild-type strain and was amplified together with the trpC terminator (TtrpC) of A. nidulans from plasmid pLUC-TargB02 (Volker Schroeckh, personal communication) using primer pair P17/P18. All obtained DNA fragments were assembled using the NEBuilder HiFi DNA Assembly Master Mix (New England Biolabs, Frankfurt, Germany) according to the manufacturer’s instructions. Transformation of A. nidulans was carried out as described before (Ballance and Turner, 1985) and transformants were selected on AMM agar plates supplemented with 3 mg/L para-aminobenzoic acid (Sigma-Aldrich, Hamburg, Germany).
For the transfer of the fgn cluster from A. fumigatus ATCC 46645 to A. nidulans RMS011, genes fgnA, Afu1g01000, Afu1g00990, Afu1g00980, Afu1g00970 were PCR-amplified using the primer pairs P19/P20, P21/P22, P23/P24, P25/P26, P27/P28, respectively. Each of the obtained DNA fragments were cloned into a single plasmid. The DNA sequences were separated by 2A sequences, as previously described (Hoefgen et al., 2018). The generated plasmid was used for transformation of A. nidulans RMS011. Transformants were selected on AMM agar plates supplemented with 3 mg/L para-aminobenzoic acid (Sigma-Aldrich, Hamburg, Germany). As a result, all genes were transcribed as a polycistronic mRNA in A. nidulans.
Southern blot analysis
Request a detailed protocolSouthern blotting was performed using a digoxigenin-11-dUTP-labeled (Jena Bioscience, Jena, Germany) probe (Schroeckh et al., 2009). The primers used for probe amplification are listed in Supplementary file 4.
Brightfield and fluorescence microscopy
Request a detailed protocolFor image acquisition of Streptomyces spores or mycelia growing in 48-well plates prepared as described above, a Keyence BZ-X800 microscope was used with 4× magnification for mycelia and 20× phase contrast magnification for imaging of spores. For fluorescence microscopy of fungal mycelia, a Zeiss Axio162 Imager M2-fluorescence microscope (Zeiss, Jena, Germany) was used. A. nidulans strains were grown on thin AMM agar plates for 16 h at 37°C, with necessary supplements (e.g. para-aminobenzoic acid and doxycycline for the tetOn inducible mutant strains, or para-aminobenzoic acid and arginine for the wild-type strain). An agar plug of 1 cm diameter was removed and placed on a microscopy slide. Nuclei were stained with Hoechst 34580 (Thermo Fisher Scientific, Dreieich, Germany) and visualized with 20× magnification.
Germination assay
Request a detailed protocolStreptomyces spores (1 × 107) were incubated in 48-well plates (Thermo Scientific Nunc, Frankfurt, Germany) in presence or absence of fumigermin or germicidin B at 28°C and shaking at 500 rpm in a Plate Shaker (PST-60HL, BioSan, Riga, Latvia). To analyze the impact of the compound on the germination, the OD595 of the spore suspension was measured every 24 h for 90 h in a Tecan fluorometer (Infinite M200 PRO, Männedorf, Switzerland). Germicidin B (Cayman Chemical, Ann Arbor, USA) and methanol were used as controls. The inhibition rate was calculated based on an OD595 measurement as previously described (Petersen et al., 1993).
Data availability
All data generated or analysed during this study are included in the manuscript. Relevant additional information has been included in the supplementary material.
References
-
Biosynthesis of silver nanoparticles by a new strain of Streptomyces sp. compared with Aspergillus fumigatusWorld Journal of Microbiology and Biotechnology 28:1081–1086.https://doi.org/10.1007/s11274-011-0906-0
-
Physiological role of germicidins in spore germination and hyphal elongation in Streptomyces coelicolor A3(2)The Journal of Antibiotics 64:607–611.https://doi.org/10.1038/ja.2011.59
-
Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulansNature Chemical Biology 3:213–217.https://doi.org/10.1038/nchembio869
-
Secondary metabolite arsenal of an opportunistic pathogenic fungusPhilosophical Transactions of the Royal Society B: Biological Sciences 371:20160023.https://doi.org/10.1098/rstb.2016.0023
-
Regulation of fungal secondary metabolismNature Reviews Microbiology 11:21–32.https://doi.org/10.1038/nrmicro2916
-
Biochemical and structural characterization of germicidin synthase: analysis of a type III polyketide synthase that employs acyl-ACP as a starter unit donorJournal of the American Chemical Society 134:7359–7366.https://doi.org/10.1021/ja2112228
-
Secondary metabolites produced during the germination of Streptomyces coelicolorFrontiers in Microbiology 8:2495.https://doi.org/10.3389/fmicb.2017.02495
-
The natural history of antibioticsCurrent Biology 19:R437–R441.https://doi.org/10.1016/j.cub.2009.04.001
-
Polyketides, proteins and genes in fungi: programmed nano-machines begin to reveal their secretsOrganic & Biomolecular Chemistry 5:2010–2026.https://doi.org/10.1039/b704420h
-
Pestalone, a new antibiotic produced by a marine fungus in response to bacterial challengeJournal of Natural Products 64:1444–1446.https://doi.org/10.1021/np0102713
-
Streptomyces viridochromogenes spore germination initiated by calcium ionsJournal of Bacteriology 143:377–382.https://doi.org/10.1128/JB.143.1.377-382.1980
-
Loss of aerial mycelium and other changes in streptomycete development due to physical variations of cultural conditionsJournal of General Microbiology 13:136–148.https://doi.org/10.1099/00221287-13-1-136
-
Interference competition among coprophilous fungi: production of (+)-Isoepoxydon by Poronia punctataApplied and Environmental Microbiology 54:861–864.https://doi.org/10.1128/AEM.54.4.861-864.1988
-
Streptomyces iranensis sp. nov., isolated from soilInternational Journal of Systematic and Evolutionary Microbiology 60:1504–1509.https://doi.org/10.1099/ijs.0.015339-0
-
Upgrading fungal gene expression on demand: improved systems for doxycycline-dependent silencing in Aspergillus fumigatusApplied and Environmental Microbiology 79:1751–1754.https://doi.org/10.1128/AEM.03626-12
-
Dispersed growth of Streptomyces in liquid cultureApplied Microbiology and Biotechnology 31:272–277.https://doi.org/10.1007/BF00258408
-
Gibepyrone biosynthesis in the rice pathogen Fusarium fujikuroi Is Facilitated by a Small Polyketide Synthase Gene ClusterJournal of Biological Chemistry 291:27403–27420.https://doi.org/10.1074/jbc.M116.753053
-
Mycobacterium smegmatis alters the production of secondary metabolites by marine-derived Aspergillus nigerJournal of Natural Medicines 74:76–82.https://doi.org/10.1007/s11418-019-01345-0
-
Fungal secondary metabolism - from biochemistry to genomicsNature Reviews Microbiology 3:937–947.https://doi.org/10.1038/nrmicro1286
-
Fungal secondary metabolism: regulation, function and drug discoveryNature Reviews Microbiology 17:167–180.https://doi.org/10.1038/s41579-018-0121-1
-
BookPractical Streptomyces GeneticsNorwich, United Kingdom: John Innnes Foundation.
-
Five new members of the Streptomyces violaceusniger 16S rRNA gene clade: Streptomyces castelarensis sp. nov., comb. nov., Streptomyces himastatinicus sp. nov., Streptomyces mordarskii sp. nov., Streptomyces rapamycinicus sp. nov. and Streptomyces ruanii sp. novInternational Journal of Systematic and Evolutionary Microbiology 58:1369–1378.https://doi.org/10.1099/ijs.0.65408-0
-
Identification of a polyketide synthase gene (pksP) of Aspergillus fumigatus involved in conidial pigment biosynthesis and virulenceMedical Microbiology and Immunology 187:79–89.https://doi.org/10.1007/s004300050077
-
Regulation and role of fungal secondary metabolitesAnnual Review of Genetics 50:371–392.https://doi.org/10.1146/annurev-genet-120215-035203
-
CDD/SPARCLE: functional classification of proteins via subfamily domain architecturesNucleic Acids Research 45:D200–D203.https://doi.org/10.1093/nar/gkw1129
-
CD-Search: protein domain annotations on the flyNucleic Acids Research 32:W327–W331.https://doi.org/10.1093/nar/gkh454
-
Imaging mass spectrometry of a coral microbe interaction with fungiJournal of Chemical Ecology 39:1045–1054.https://doi.org/10.1007/s10886-013-0320-1
-
Microbial interactions trigger the production of antibioticsCurrent Opinion in Microbiology 45:117–123.https://doi.org/10.1016/j.mib.2018.04.002
-
Genome-Based taxonomic classification of the phylum ActinobacteriaFrontiers in Microbiology 9:2007.https://doi.org/10.3389/fmicb.2018.02007
-
Germicidin, an autoregulative germination inhibitor of Streptomyces viridochromogenes NRRL B-1551The Journal of Antibiotics 46:1126–1138.https://doi.org/10.7164/antibiotics.46.1126
-
Phagensensibilität, Zellwand-Zusammensetzung und taxonomie von actinomycetenJournal of Basic Microbiology 8:39–46.https://doi.org/10.1002/jobm.19680080106
-
Discovery of microbial natural products by activation of silent biosynthetic gene clustersNature Reviews Microbiology 13:509–523.https://doi.org/10.1038/nrmicro3496
-
Methods for characterization of Streptomyces species1International Journal of Systematic and Evolutionary Microbiology 16:313.https://doi.org/10.1099/00207713-16-3-313
-
Type III polyketide synthase beta-ketoacyl-ACP starter unit and ethylmalonyl-CoA extender unit selectivity discovered by Streptomyces coelicolor genome miningJournal of the American Chemical Society 128:14754–14755.https://doi.org/10.1021/ja065247w
-
Rodletless, a new Aspergillus developmental mutant induced by directed gene inactivationGenes & Development 5:1161–1171.https://doi.org/10.1101/gad.5.7.1161
-
Fusion PCR and gene targeting in Aspergillus nidulansNature Protocols 1:3111–3120.https://doi.org/10.1038/nprot.2006.405
-
Host and virus determinants of Picornavirus pathogenesis and tropismNature Reviews Microbiology 3:765–776.https://doi.org/10.1038/nrmicro1284
Article and author information
Author details
Maria Cristina Stroe

Funding
Deutsche Forschungsgemeinschaft (Collaborative Research Center 1127 ChemBioSys (projects B01))
- Christian Hertweck
Deutsche Forschungsgemeinschaft (Collaborative Research Center 1127 ChemBioSys (projects B02))
- Tina Netzker
- Axel A Brakhage
Bundesministerium für Bildung und Forschung (DrugBioTune)
- Maria Cristina Stroe
- Axel A Brakhage
International Leibniz Research School (Microbial and Biomolecular Interactions)
- Axel A Brakhage
- Maria Cristina Stroe
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Christiane Weigel for antimicrobial assays, Carmen Schult, Silke Steinbach and Karin Burmeister for excellent technical assistance, Heike Heinecke for NMR measurements, and Sven Krappmann and Volker Schroeckh for their kind plasmid donations. Financial support by the International Leibniz Research School for Microbial and Biomolecular Interactions (ILRS), the Deutsche Forschungsgemeinschaft (DFG)-funded Collaborative Research Center 1127 ChemBioSys (projects B01, B02) and the Bundesministerium für Bildung und Forschung (BMBF)-funded project DrugBioTune in the frame of InfectControl2020 is gratefully acknowledged.
Copyright
© 2020, Stroe et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 4,577
- views
-
- 858
- downloads
-
- 77
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 77
- citations for umbrella DOI https://doi.org/10.7554/eLife.52541
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Targeted induction of a silent fungal gene cluster encoding the bacteria-specific germination inhibitor fumigermin
eLife 9:e52541.
https://doi.org/10.7554/eLife.52541




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}