Oxytocin signaling in the posterior hypothalamus prevents hyperphagic obesity in mice
- RIKEN Center for Biosystems Dynamics Research, Japan
- Laboratory of Molecular Biology, Department of Molecular and Cell Biology, Graduate School of Agricultural Science, Tohoku University, Japan
- Division of Brain and Neurophysiology, Department of Physiology, Jichi Medical University, Japan
- Department of Obesity and Inflammation Research, Fukushima Medical University, Japan
- CREST, Japan Science and Technology Agency, Japan
Abstract
Decades of studies have revealed molecular and neural circuit bases for body weight homeostasis. Neural hormone oxytocin (Oxt) has received attention in this context because it is produced by neurons in the paraventricular hypothalamic nucleus (PVH), a known output center of hypothalamic regulation of appetite. Oxt has an anorexigenic effect, as shown in human studies, and can mediate satiety signals in rodents. However, the function of Oxt signaling in the physiological regulation of appetite has remained in question, because whole-body knockout (KO) of Oxt or Oxt receptor (Oxtr) has little effect on food intake. We herein show that acute conditional KO (cKO) of Oxt selectively in the adult PVH, but not in the supraoptic nucleus, markedly increases body weight and food intake, with an elevated level of plasma triglyceride and leptin. Intraperitoneal administration of Oxt rescues the hyperphagic phenotype of the PVH Oxt cKO model. Furthermore, we show that cKO of Oxtr selectively in the posterior hypothalamic regions, especially the arcuate hypothalamic nucleus, a primary center for appetite regulations, phenocopies hyperphagic obesity. Collectively, these data reveal that Oxt signaling in the arcuate nucleus suppresses excessive food intake.
Editor's evaluation
Inada and colleagues report results from a series of studies investigating the role of the neuropeptide oxytocin in regulating food intake and body weight. They used combinations of genetics and behavioral studies to demonstrate that oxytocin deletion results in increased food intake and body weight. They further show that deletion of the oxytocin receptor in the posterior hypothalamus causes a similar increase in food intake and body weight. Collectively, these studies support a role for the oxytocin system as a key regulator of energy balance.
https://doi.org/10.7554/eLife.75718.sa0Introduction
Appetite is one of the strongest desires in animals. The consumption of nutritious foods is a primitive pleasure for animals because it is essential for survival. Yet, excessive food intake leads to obesity and increases the risk of disease. Understanding the neurobiological bases of appetite regulation is therefore an urgent issue, given that the body mass index of humans has increased dramatically over the last 40 years (NCD Risk Factor Collaboration, 2016).
Decades of studies in rodents have revealed molecular and neural circuit bases for body weight homeostasis (Andermann and Lowell, 2017; Sternson and Eiselt, 2017; Sutton et al., 2016). Classical studies with mechanical or electrical lesioning, as well as recent molecular or genetic dissections, both support the critical roles of appetite regulation by neurons in the arcuate hypothalamic nucleus (ARH), in particular, those expressing orexigenic agouti-related protein (Agrp) and anorexigenic pre-opiomelanocortin (Pomc) (Choi et al., 1999; Ollmann et al., 1997). These neurons receive both direct humoral inputs and neural inputs of interoception (Bai et al., 2019) to regulate food intake antagonistically at various timescales (Krashes et al., 2014; Sternson and Eiselt, 2017). The paraventricular hypothalamic nucleus (PVH) is one of the critical output structures of the primary appetite-regulating ARH neurons. Silencing PVH neurons phenocopies the overeating effect observed in the activation of Agrp neurons, whereas activating PVH neurons ameliorates the overeating caused by the acute activation of Agrp neurons (Atasoy et al., 2012; Garfield et al., 2015). Melanocortin-4 receptor (MC4R)-expressing neurons in the PVH are the key target of ARH Agrp and Pomc neurons. PVH MC4R neurons are activated by α-melanocyte-stimulating hormone provided by Pomc neurons and inhibited by GABAergic Agrp neurons, and are supposed to transmit signals to the downstream target regions in the midbrain and pons (Garfield et al., 2015; Stachniak et al., 2014; Sutton et al., 2016).
Although PVH MC4R neurons have been relatively well documented (Balthasar et al., 2005; Garfield et al., 2015), other PVH cell types may also mediate output signals to control feeding and energy expenditure (Sutton et al., 2016). However, little is known about the organization, cell types, and neurotransmitters by which appetite-regulating signals are conveyed to other brain regions. Neural hormone oxytocin (Oxt), which marks one of the major cell types in the PVH, has received attention in this context (Leng and Sabatier, 2017; Onaka and Takayanagi, 2019). The anorexigenic effect of Oxt has been shown in humans (Lawson et al., 2015; Thienel et al., 2016), and genetic variations of Oxt receptor (Oxtr) have been implicated as a risk factor of overeating (Çatli et al., 2021; Davis et al., 2017). In rodents, Oxt administration has been shown to suppress increases in food intake and body weight (Maejima et al., 2018). Pons-projecting Oxt neurons have been shown to be active following leptin administration (Blevins et al., 2004), and knockdown of Oxtr in the nucleus of the solitary tract has been reported to alter feeding patterns (Ong et al., 2017). In addition, Oxtr-expressing neurons in the ARH have been shown to evoke acute appetite suppression signals when opto- or chemo-genetically activated (Fenselau et al., 2017). Despite the importance of the Oxt-OxtR system in the food intake and homeostasis of body weight (Maejima et al., 2018), knockout (KO) and ablation studies still question such findings (Sutton et al., 2016; Worth and Luckman, 2021). For example, Oxt or Oxtr KO mice showed increased body weight at around 4 months of age (termed late-onset obesity), while their food intake was not different from that of wild-type mice (Camerino, 2009; Takayanagi et al., 2008). Diphtheria toxin-based genetic ablation of Oxt-expressing cells in adult mice increased the body weight of male mice with a high-fat diet, but not those with normal chow, and in both cases, food intake was unaffected (Wu et al., 2012b). To revisit the function and sites of action of Oxt signaling in the regulation of feeding, acute conditional KO (cKO) mouse models would be useful.
Here, we describe Oxt cKO phenotypes related to hyperphagic obesity. Our approach offers the following two advantages over previous studies: (i) the Oxt gene can be knocked out in adult mice, avoiding the influence of possible developmental and genetic compensations (El-Brolosy et al., 2019); and (ii) the manipulation can be restricted to the brain, or even to a single hypothalamic nucleus, providing a resolution that exceeds previous studies. Owing to these advantages, we show that Oxt cKO increases both body weight and food intake. The suppression of overeating and overweighting is predominantly regulated by Oxt neurons in the PVH, leaving Oxt neurons in the supraoptic nucleus (SO) with only a minor role. We further show that Oxtr-expressing neurons in the posterior part of the hypothalamus, especially the ARH, mediate the overeating-suppression signals generated by Oxt neurons.
Results
cKO of PVH Oxt increases body weight and food intake
To examine the necessity of Oxt for the regulation of food intake, we prepared recently validated Oxtflox/flox mice (Inada et al., 2022). In this line, Cre expression deletes floxed exon 1 of the Oxt gene, resulting in the loss of transcription of Oxt mRNA (Inada et al., 2022). To perform the cKO in PVH Oxt neurons, we first crossed Oxtflox/flox and Oxt KO (Oxt–/–) mice and obtained Oxtflox/– mice. Then, we injected AAV-Cre into the bilateral PVH of 8-week-old Oxtflox/– male mice (Figure 1A and B). The number of neurons expressing Oxt, visualized by in situ hybridization (ISH), significantly decreased within 3 weeks after the AAV-Cre injection (Figure 1C). The body weight of Oxtflox/– mice that received AAV-Cre injection started to deviate from the controls at around 3 weeks after the injection (Figure 1D). At 5 weeks after the injection, we compared the body weight of Oxtflox/– mice that received AAV-Cre injection with the wild-type (Oxt+/+), Oxt–/–, and Oxtflox/– mice that received vehicle injection. We found that AAV-Cre-injected Oxtflox/– mice were heavier than those in the other groups (Figure 1E and F). Importantly, this increase in body weight was considered unlikely to be a reflection of late-onset obesity, as previously reported (Camerino, 2009; Takayanagi et al., 2008), because we did not find a significant difference between the wild-type and Oxt–/– mice (Figure 1F). Next, we analyzed the relationship between the number of remaining Oxt+ neurons and body weight. We found that mice with a fewer number of remaining Oxt+ neurons showed a heavier body weight (Figure 1G). We also found an increase in food intake: the daily food intake of Oxtflox/– mice that received AAV-Cre injection was significantly larger at >4 weeks after the injection (Figure 1H), and the total food intake during the 5 weeks after the injection was also larger in the mice that received AAV-Cre injection (Figure 1I). Of note, these effects are not due to the nonspecific toxicity of AAV-Cre injection per se, as AAV-Cre injection to wild-type mice did not alter body weight or daily food intake (Figure 1—figure supplement 1A–D). These results demonstrate that the cKO of Oxt evokes increases in both body weight and food intake.
Figure 1 with 1 supplement see all
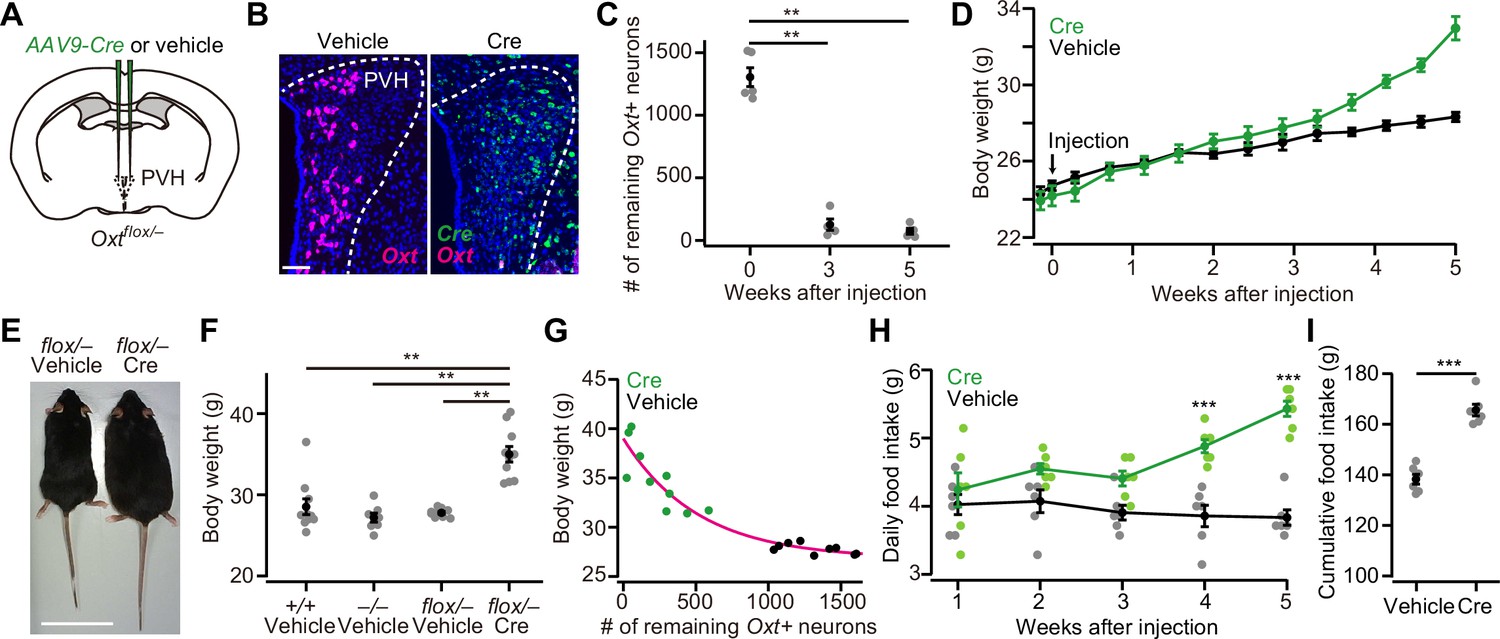
Oxytocin (Oxt) conditional knockout (cKO) in paraventricular hypothalamic nucleus (PVH) induces an increase in body weight and food intake.
(A) Schematic of the virus injection. AAV-Cre or vehicle was injected into the bilateral PVH of Oxtflox/– male mice. (B) Representative coronal sections of the PVH from Oxtflox/– mice received vehicle (left) or AAV-Cre (right) injection. Data were obtained at 5 weeks after the injection. Magenta and green represent Oxt and Cre in situ stainings, respectively. Blue, DAPI. Scale bar, 50 μm. (C) The number of remaining Oxt+ neurons in the PVH of mice that received AAV-Cre injection. **p<0.01, one-way ANOVA with post hoc Tukey’s HSD. N=5 each. (D) Time course of body weight after AAV-Cre or vehicle injection. N=6 each. (E) Representative photos of Oxtflox/– mice that received either vehicle (left) or AAV-Cre injection (right). Five weeks after the injection. Scale bar, 5 cm. (F) Body weight of wild-type (+/+), Oxt KO (–/–), and Oxt cKO (flox/–) mice. The weight was measured at 5 weeks after injection of either vehicle or AAV-Cre. Note that this time point corresponds to 13 weeks of age. **p<0.01, one-way ANOVA with post hoc Tukey’s HSD. N=10, 7, 9, and 10 for +/+, –/–, flox/– vehicle, and flox/– Cre, respectively. (G) Relationship between the number of remaining Oxt+ neurons in the PVH and the body weight of Oxtflox/– mice shown in (F). Magenta, exponential fit for the data from both Cre and vehicle. (H) Time course of daily food intake, defined as the average food intake in each week after AAV-Cre or vehicle injection. ***p<0.001, Student’s t-test with post hoc Bonferroni correction. N=6 each. (I) Cumulative food intake during the 5 weeks after the injection. ***p<0.001, Student’s t-test. N=6 each.
Error bars, standard error of mean (SEM).
In addition to the PVH, Oxt neurons are also clustered in the SO (Zhang et al., 2021). To examine whether Oxt neurons in the SO also play inhibitory roles on body weight and food intake, we injected AAV-Cre into the bilateral SO of Oxtflox/– mice (Figure 2A and B). Similar to the PVH, the number of Oxt+ neurons in the SO significantly decreased at around 3 weeks after the injection (Figure 2C). Unlike the PVH, however, neither body weight nor food intake was significantly different compared with controls (Figure 2D–G), and no clear relationship was found between the number of the remaining Oxt+ neurons and body weight (Figure 2E). These results suggest that PVH Oxt neurons predominantly regulate food intake and body weight, and that SO Oxt neurons exert little influence.
Figure 2
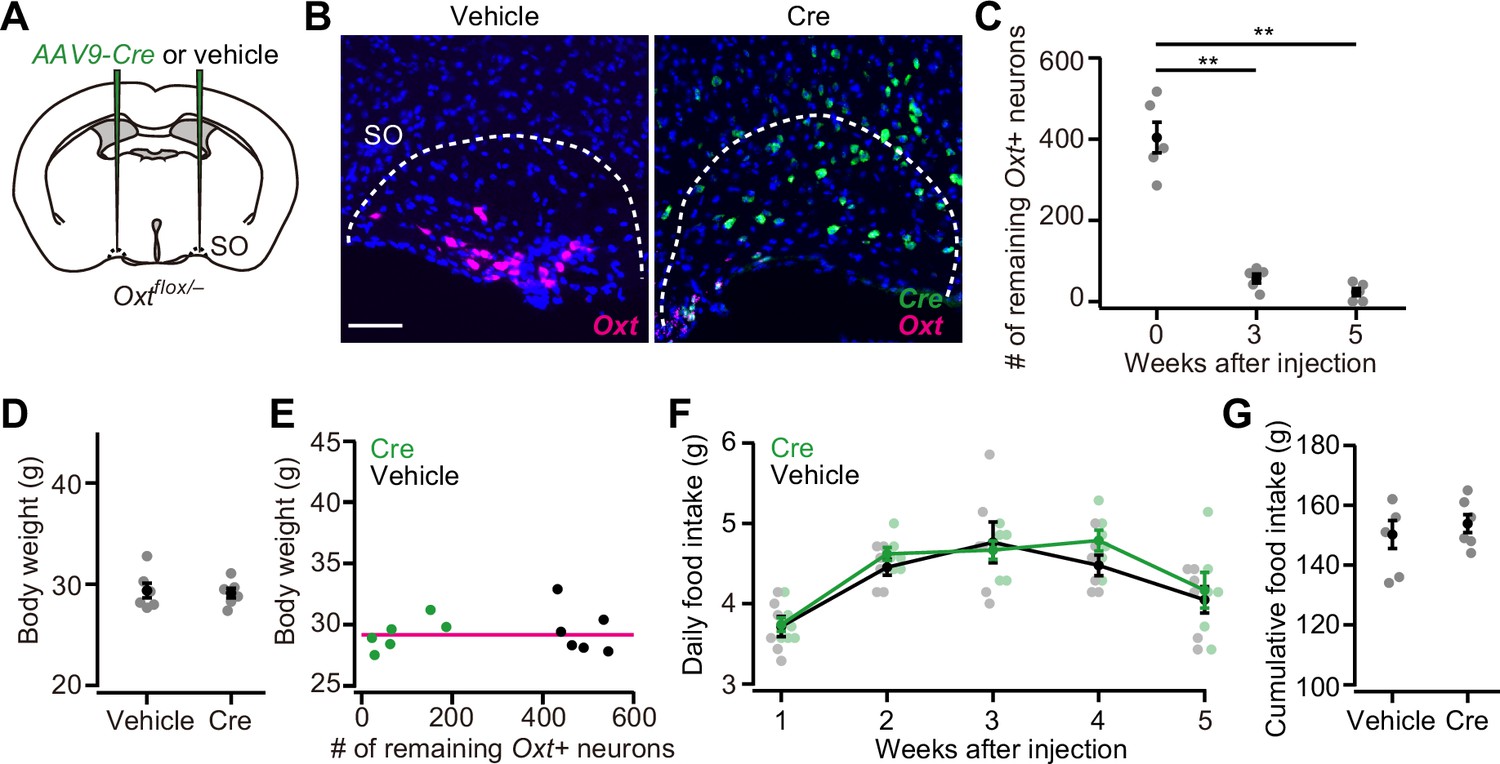
Oxytocin (Oxt) conditional knockout (cKO) in supraoptic nucleus (SO) has a negligible effect on food intake and body weight.
(A) Schematic of the virus injection. AAV-Cre or vehicle was injected into the bilateral SO of Oxtflox/– male mice. (B) Representative coronal sections of left SO from Oxtflox/– mice received vehicle (left) or AAV-Cre (right) injection. Five weeks after the injection. Magenta and green represent Oxt and Cre in situ stainings, respectively. Blue, DAPI. Scale bar, 50 μm. (C) The number of remaining Oxt+ neurons in the SO of mice that received AAV-Cre injection. **p<0.01, one-way ANOVA with post hoc Tukey’s HSD. N=5 each. (D) The body weight of Oxtflox/– mice did not differ between vehicle or AAV-Cre (Student’s t-test). N=6 each. Data were obtained at 5 weeks after the injection. (E) Relationship between the number of remaining Oxt+ neurons in the SO and the body weight of Oxtflox/– mice shown in (D). Magenta, exponential fit for the data from both Cre and vehicle. (F) The time course of daily food intake was not statistically different (Student’s t-test with post hoc Bonferroni correction). N=6 each. (G) Cumulative food intake during the 5 weeks after the injection. N=6 each.
Error bars, SEM.
Because of the minor role of SO Oxt neurons, we focused on the PVH Oxt neurons in the following experiments.
Weight of viscera and blood constituents
Increased food intake may influence not only body weight, but also the viscera and blood constituents. To examine these points, we collected internal organs and blood samples from non-fasted Oxtflox/– mice that had received either AAV-Cre or vehicle injection into the bilateral PVH (Figure 3A). While the weight of the stomach was unchanged (Figure 3B), a significant increase was observed in the weight of the liver in Oxtflox/– mice with AAV-Cre injection, likely because of the accumulation of fat in the liver (Figure 3C). We next measured the plasma concentration of glucose, triglyceride, and leptin. No significant differences in glucose levels were found (Figure 3D). In turn, the plasma concentrations of triglyceride and leptin were higher in Oxtflox/– mice that had received AAV-Cre injection than in those that had received vehicle injection (Figure 3D). Of note, a prominent increase in plasma leptin was also reported in the late-onset obesity cases of 6-month-old Oxt KO mice (Camerino, 2009). Our data regarding Oxt cKO showed the plasma leptin phenotype in the earlier stage of 13-week-old mice. These results suggest that the cKO of Oxt affects the homeostasis of viscera and blood constituents.
Figure 3

Weight of viscera and the blood constituents.
(A) Schematic of the virus injection. AAV-Cre or vehicle was injected into the bilateral paraventricular hypothalamic nucleus (PVH) of Oxtflox/– male. Data were obtained at 5 weeks after the injection. (B) The weight of the stomach was not statistically different (p>0.5, Student’s t-test. N=9 and 10 for vehicle and Cre, respectively). (C) The weight of the liver was significantly heavier in AAV-Cre-injected mice (***p<0.001, Student’s t-test). (D) Plasma glucose (left), triglyceride (middle), and leptin (right) measured in the non-fasted Oxtflox/– mice. **p<0.01, ***p<0.001, Student’s t-test. N=9 and 8 mice for vehicle and Cre, respectively.
Error bars, SEM.
Oxt supplementation partially rescues Oxt cKO
If Oxt cKO caused increases in body weight and food intake with higher plasma triglyceride and leptin, such effects might be mitigated by the external administration of Oxt. This hypothesis is also supported by the fact that intraperitoneal (ip) or intracerebroventricular injection of Oxt has been shown to reduce both body weight and food intake (Maejima et al., 2018). To examine this hypothesis, first, we injected AAV-Cre into the bilateral PVH of Oxtflox/– mice, and from the next day of the injection, we conducted ip injection of Oxt (100 μL of 500 μM or 1 mM solution) once every 3 days (Figure 4A). Ip injection of the vehicle was used as control. At 5 weeks after the injection of AAV-Cre, Oxt-treated mice showed significantly reduced body weight, even though the number of remaining Oxt+ neurons was not significantly different (Figure 4B and C). We found that both daily food intake at 4–5 weeks and total food intake during the 5 weeks after the injection were also significantly reduced (Figure 4D and E). The reduction of body weight and food intake by our Oxt treatment paradigm was somehow specific to Oxt cKO mice, given that neither reduction of food intake nor body weight was observed in wild-type males (Figure 4—figure supplement 1A–D). No significant improvement in the blood samples was found: both plasma triglyceride and leptin tended to be reduced in Oxt-treated mice, but did not reach the level of statistical significance (Figure 4F). These results suggest that external administration of Oxt can rescue at least the hyperphagic obesity phenotype of Oxt cKO.
Figure 4 with 1 supplement see all
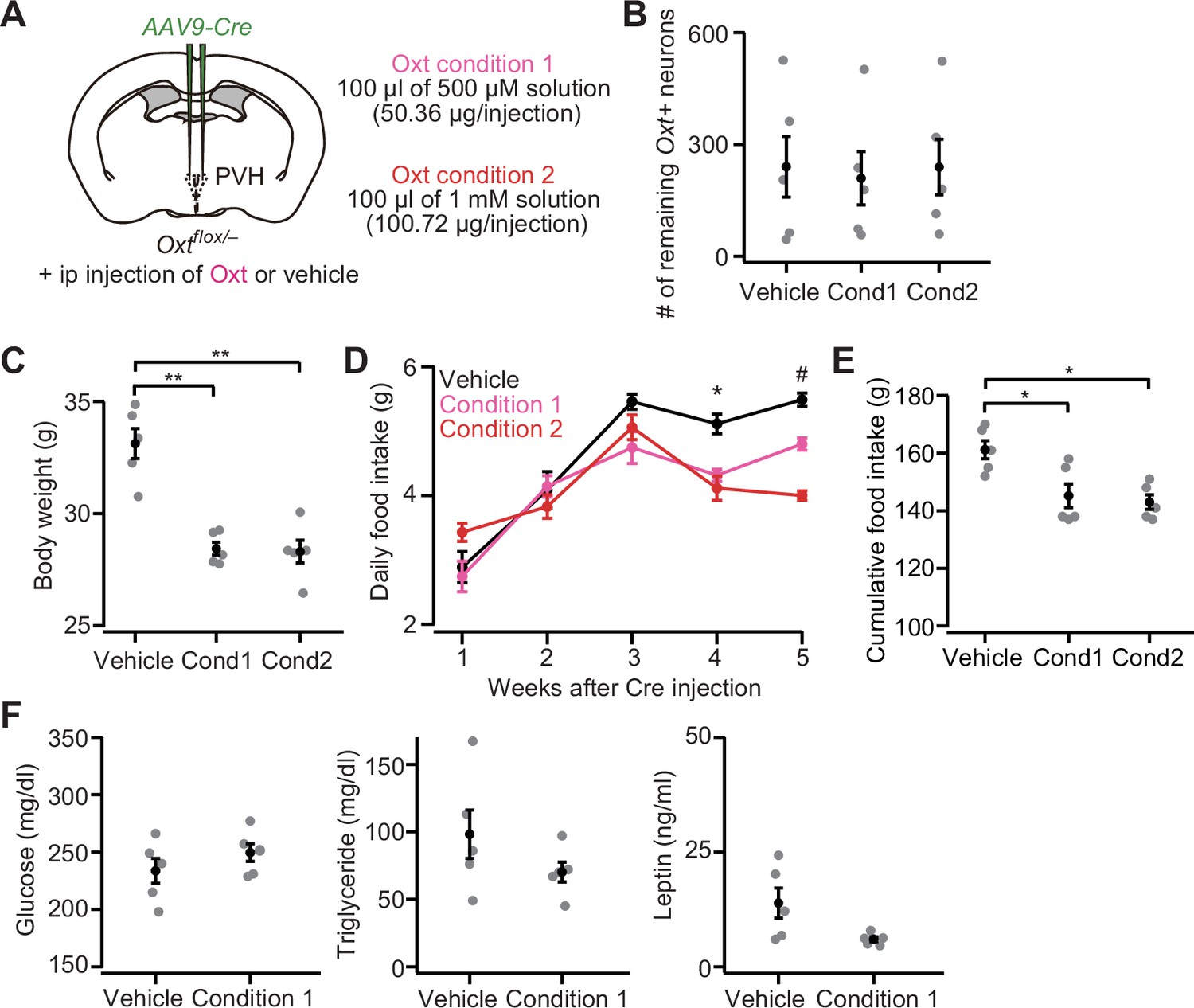
Intraperitoneal (ip) injection of oxytocin (Oxt) partially rescues paraventricular hypothalamic nucleus (PVH) Oxt conditional knockout (cKO) phenotypes.
(A) Schematic of the experiments. AAV-Cre was injected into the bilateral PVH of Oxtflox/– male mice. Data were obtained 5 weeks after the virus injection. Once every 3 days, the mice received ip injection of vehicle, 50.36 μg of Oxt (condition 1) or 100.72 μg of Oxt (condition 2) (see Materials and methods). (B) The number of remaining Oxt+ neurons was not statistically different (p>0.9, one-way ANOVA). Cond, condition. N=5 each (same mice across panels B–E). (C) Ip injection of Oxt significantly decreased body weight. **p<0.01, one-way ANOVA with post hoc Tukey’s HSD. Cond, condition. (D) Time course of daily food intake. Asterisks (*) denote significant differences for vehicle versus condition 1 and vehicle versus condition 2 (p<0.05, Tukey’s HSD), and hashes (#) denote significant differences for vehicle versus condition 1, vehicle versus condition 2, and condition 1 versus condition 2 (p<0.05, Tukey’s HSD). (E) Cumulative food intake during the 5 weeks after the virus injection was decreased in the mice that received ip injection of Oxt (*p<0.05, one-way ANOVA with post hoc Tukey’s HSD). Cond, condition. (F) Plasma glucose (left), triglyceride (middle), and leptin (right) measured in the non-fasted Oxtflox/– mice. Decreases in triglyceride and leptin on average were found in Oxt-treated mice but did not reach the level of statistical significance (p=0.314 and 0.065 for triglyceride and leptin, respectively, Student’s t-test). N=5 each.
Error bars, SEM.
In addition to the daily food intake that we have examined so far, previous studies showed that mice ate less within several hours after receiving ip injection of Oxt (Arletti et al., 1989; Maejima et al., 2011). To examine whether Oxtflox/– mice that receive AAV-Cre injection similarly show reduced hourly food intake, we measured food intake after 6 hr of fasting (Figure 4—figure supplement 1E and F). After fasting, the mice received an ip injection of Oxt, and food was provided again (Figure 4—figure supplement 1F). Cumulative food intake was measured at 1, 3, and 5 hr after the placement of food (Figure 4—figure supplement 1F). Although the number of remaining Oxt+ neurons was comparable (Figure 4—figure supplement 1G), Oxt-injected mice ate less (Figure 4—figure supplement 1H). Taken together, ip injection of Oxt appears to reduce food intake on the scale of hours to days, thereby preventing the hyperphagic obesity induced by Oxt cKO in the PVH.
Oxtr-expressing cells in the ARH mediate appetite suppression
Having established the importance of Oxt to suppress hyperphagic obesity, we examined the site of action of Oxt signaling that mediates appetite suppression. To this end, we prepared Oxtrflox/flox mice, in which the Oxtr gene can be knocked out under Cre expression (Takayanagi et al., 2005). Given that PVH Oxt neurons send dense projection to the hypothalamic nuclei (Yao et al., 2017; Zhang et al., 2021), and that OxtR expression is also found in the hypothalamus (Fenselau et al., 2017; Mitre et al., 2016; Newmaster et al., 2020), we suspected that a fraction of appetite suppression signals is mediated by the other nuclei of the hypothalamus. To test this possibility, we injected AAV-Cre (serotype 9) into the bilateral ‘anterior hypothalamus’, mainly aiming at nuclei such as the anteroventral periventricular nucleus, medial preoptic nucleus medial part (MPNm), MPN lateral part, and medial preoptic area (Figure 5A; see Materials and methods), and the ‘posterior hypothalamus’, containing nuclei such as the dorsomedial nucleus of the hypothalamus, ventromedial hypothalamic nucleus (VMH), lateral hypothalamic area (LHA), and ARH (Figure 5B). AAV-mediated Cre expression roughly covered these nuclei (Figure 5C and D). To examine if Cre expression reduced Oxtr expression, we visualized Oxtr mRNA using the RNAscope assay (see the Materials and methods) (Sato et al., 2020; Wang et al., 2012). As Oxtr expression was observed as a dot-like structure (Figure 5E and F), we counted the number of such RNAscope dots in each DAPI+ cell. In a negative control experiment utilizing Oxtr KO mice, we often detected one or two RNAscope dots in the DAPI+ cells (Figure 5—figure supplement 1A and B). Therefore, we regarded a cell with three or more dots as an Oxtr-expressing cell (Oxtr+; Figure 5G and H, and Figure 5—figure supplement 2A–C). We found that AAV-Cre injection successfully reduced the number of Oxtr+ cells in most of the targeted nuclei (Figure 5G and H). Oxtr cKO in the posterior but not anterior hypothalamus significantly increased body weight (Figure 5I and J). Similarly, a significant increase in food intake was observed in the mice that had received AAV-Cre injection into the posterior hypothalamus (Figure 5K and L).
Figure 5 with 2 supplements see all
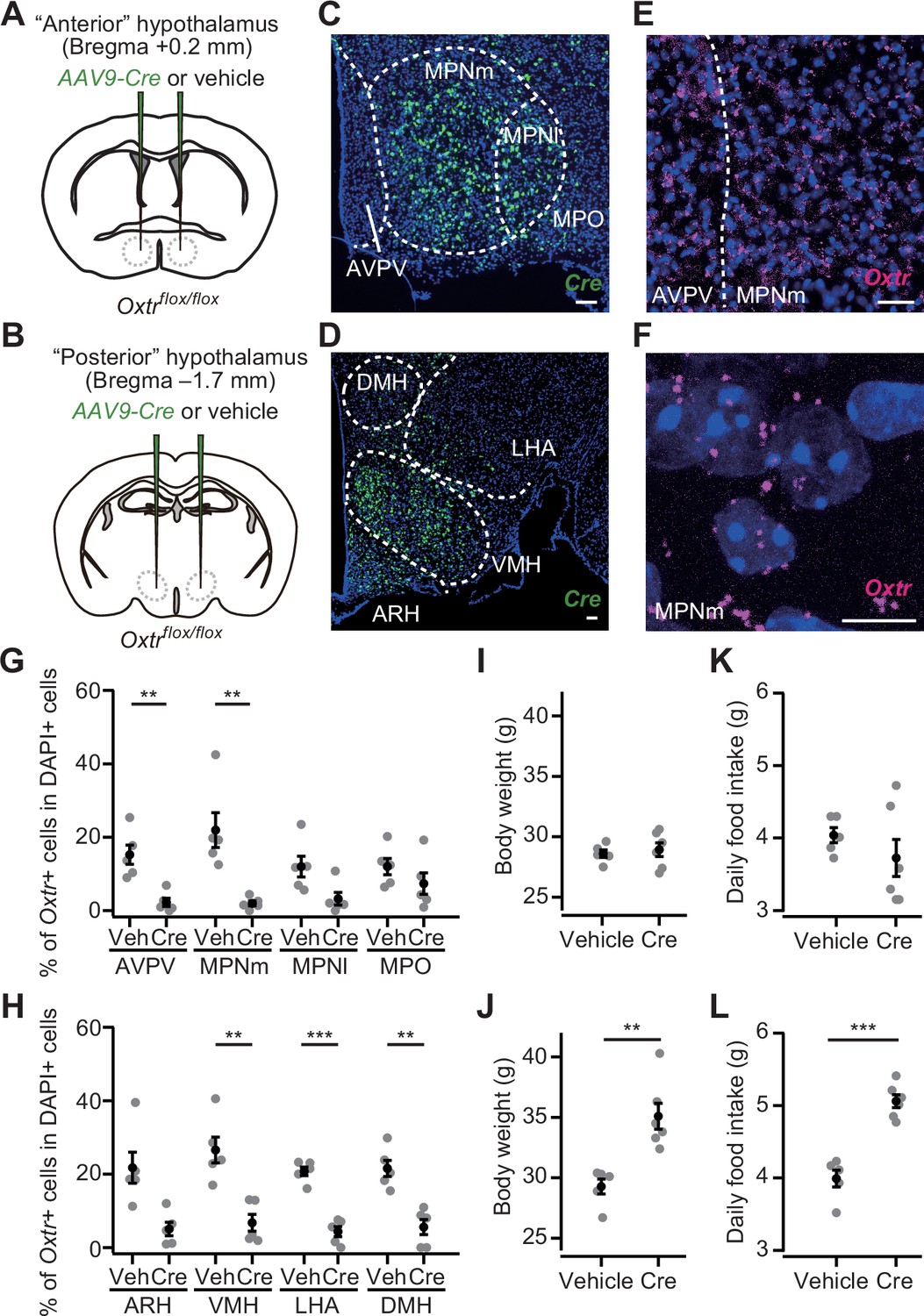
Oxtr conditional knockout (cKO) in the posterior hypothalamus induces increases in body weight and food intake.
(A, B) Schematic of the virus injection. AAV-Cre or vehicle was injected into the bilateral anterior or posterior hypothalamus (see Materials and methods) of Oxtrflox/flox male mice. (C, D) Representative coronal section showing Cre mRNA (green). Blue, DAPI. Scale bar, 50 μm. (E) A representative coronal section showing anteroventral periventricular nucleus (AVPV) and medial preoptic nucleus medial part (MPNm) from a vehicle-injected mouse. Oxtr mRNA was visualized by RNAscope (magenta). Blue, DAPI. Scale bar, 30 μm. (F) Projection of a confocal stack in MPNm from a vehicle-injected mouse. Magenta, Oxtr mRNA. Blue, DAPI. Scale bar, 5 μm. (G, H) Fraction of DAPI+ cells expressing Oxtr. Cells showing three or more RNAscope dots were defined as Oxtr+ (Figure 5—figure supplement 2). Veh, vehicle. N=5 each. **p<0.01, ***p<0.001, Student’s t-test with Bonferroni correction. Decreases in the MPN lateral part (MPNl), medial preoptic area (MPO), and arcuate hypothalamic nucleus (ARH) on average were found in AAV-Cre-injected mice but did not reach the level of statistical significance in Student’s t-test with Bonferroni correction (p=0.045, 0.289, and 0.012, respectively). (I, J) Body weight measured at 5 weeks after the injection. **p<0.01, Student’s t-test. Anterior hypothalamus, N=5 and 6 for vehicle and Cre, respectively, and posterior hypothalamus, N=5 and 6 for vehicle and Cre, respectively. (K, L) Daily food intake measured at 5 weeks after the injection. ***p<0.001, Student’s t-test. Anterior hypothalamus, N=5 and 6 for vehicle and Cre, respectively, and posterior hypothalamus, N=5 and 6 for vehicle and Cre, respectively. Error bars, SEM.
We next aimed to pinpoint a specific nucleus in the posterior hypothalamus that could suppress hyperphagic obesity. To this end, we injected AAV-Cre (serotype 2) into the ARH or LHA (Figure 6A and B). This serotype of AAV-driven Cre expression was spatially localized (Figure 6C and D) compared with the AAV serotype 9 used in Figure 5. AAV-driven Cre expression reduced Oxtr expression in Oxtrflox/flox mice (Figure 6E and F). Body weight, daily food intake, and cumulative food intake were significantly greater in the mice that received AAV-Cre injection into the ARH, whereas no significant difference was found in the mice that expressed Cre in the LHA (Figure 6G-L).
Figure 6
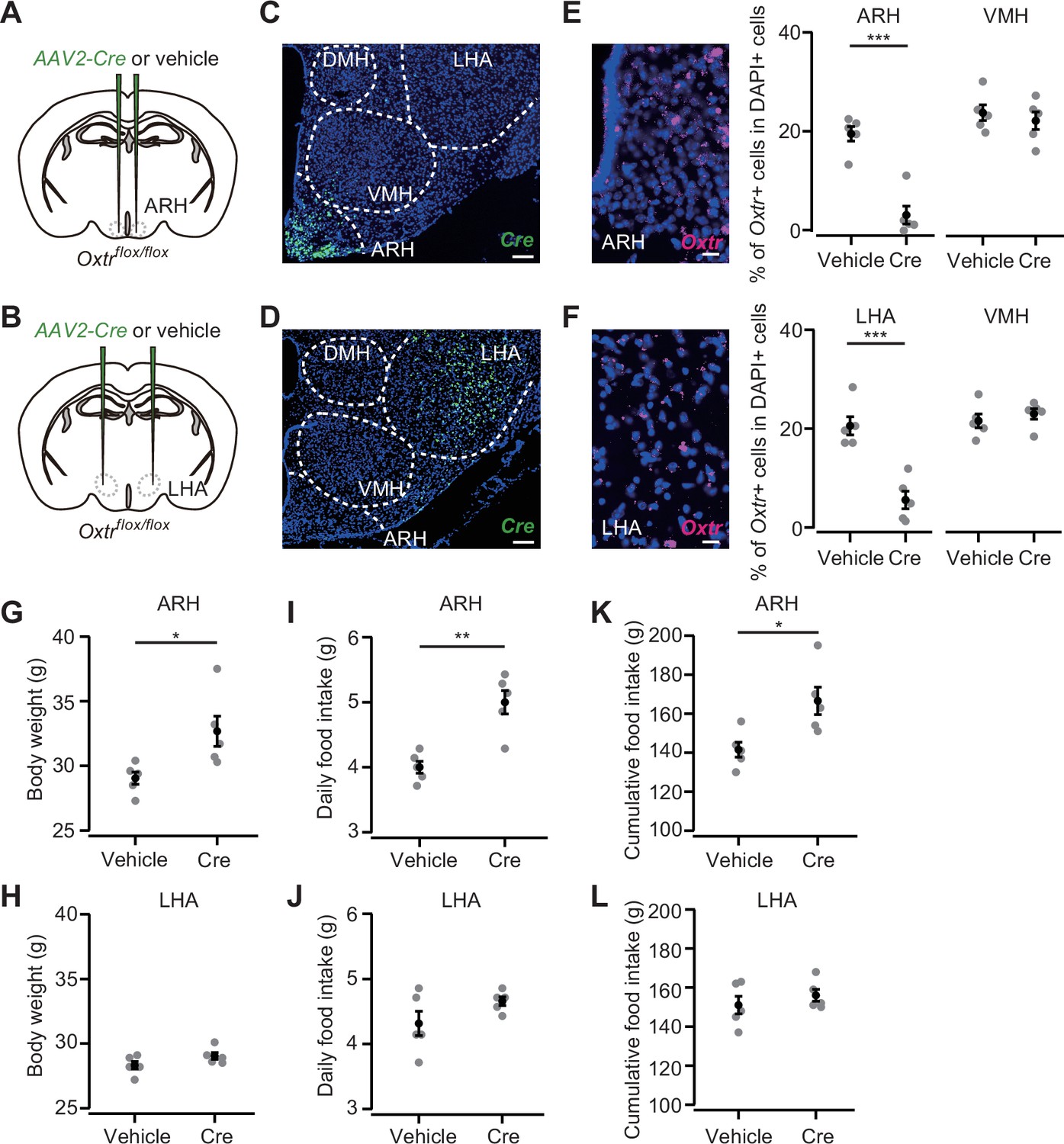
Oxytocin receptor (Oxtr) expression in the arcuate hypothalamic nucleus (ARH) suppresses body weight and food intake.
(A, B) Schematic of the virus injection. AAV-Cre or vehicle was injected into the bilateral ARH or lateral hypothalamic area (LHA) of Oxtrflox/flox male mice. (C, D) Representative coronal section showing Cre mRNA (green). Blue, DAPI. Scale bar, 50 μm. (E, F) Left, representative coronal section showing the ARH or LHA from a vehicle-injected mouse. Oxtr mRNA was visualized by RNAscope (magenta). Blue, DAPI. Scale bar, 5 μm. Right, fraction of DAPI+ cells expressing Oxtr in the ARH (E) or LHA (F) and ventromedial hypothalamic nucleus (VMH), a neighboring nucleus of the ARH and LHA. Cells showing three or more RNAscope dots were defined as Oxtr+. N=5 each. ***p<0.001, Student’s t-test with Bonferroni correction. (G, H) Body weight measured at 5 weeks after the injection. *p<0.05, Student’s t-test. N=5 each. (I, J) Daily food intake measured at 5 weeks after the injection. **p<0.01, Student’s t-test. N=5 each. (K, L) Cumulative food intake during the 5 weeks after the injection. *p<0.05, Student’s t-test. N=5 each.
Error bars, SEM.
Taken together, these results indicate that a fraction of the appetite suppression signals from Oxt neurons is mediated by Oxtr-expressing cells in the posterior hypothalamus, especially those in the ARH.
Discussion
Oxt cKO increased body weight and food intake
In this study, we performed cKO of the Oxt gene by injecting AAV-Cre, which enabled region-specific KO of Oxt. By this advantage, we showed that Oxt produced by PVH Oxt neurons contributes to the regulation of body weight and food intake, whereas that by SO Oxt neurons does not (Figures 1 and 2). These data extend the previous results that mechanical disruption of PVH in rats increased both body weight and food intake (Shor-Posner et al., 1985; Sims and Lorden, 1986). In contrast to the Oxt cKO phenotype (Figure 1H and I), whole-body Oxt KO mice showed a normal amount of food intake (Camerino, 2009), suggesting compensational mechanisms. For example, when a certain gene is knocked out, expression of the related gene(s) is enhanced to compensate for some of the KO phenotypes functionally (El-Brolosy et al., 2019; Ma et al., 2019). Transcriptomic analysis between Oxt KO and wild type may reveal a more complete picture of gene expression that can explain the compensational mechanisms in Oxt KO mice.
The phenotypic discrepancy between our data and diphtheria toxin-induced ablation of Oxt cells (Wu et al., 2012b) might be due to the loss of Oxt cells outside the PVH (maybe even outside the brain; Paiva et al., 2021) that somehow elicited appetite suppression, and therefore counterbalanced the overeating phenotype caused by the loss of Oxt in the PVH. Alternatively, neuropeptides or neurotransmitters other than Oxt expressed in the PVH Oxt neurons might have conveyed appetite-stimulating signals, which remained intact in our Oxt-selective cKO model, but were disrupted in the cell-based ablation, resulting in only the overeating phenotype to appear in our case. Regardless of the scenario, our data establish the necessity of Oxt in the PVH to suppress overeating and suggest the presence of a hormone-based output pathway of PVH appetite regulation signals, in addition to the well-established neural pathways mediated by MC4R neurons (Garfield et al., 2015; Stachniak et al., 2014; Sutton et al., 2016).
Downstream of Oxt neurons that mediate appetite suppression signals
After eating a sufficient amount of food, animals stop eating owing to the appetite suppression signals mediated in the brain. Several brain regions and cell types, such as Pomc-expressing neurons in the ARH (Ollmann et al., 1997; Sternson and Eiselt, 2017; Sutton et al., 2016), glutamatergic Oxtr-expressing neurons in the ARH (Fenselau et al., 2017), and calcitonin gene-related peptide expressing neurons in the parabrachial nucleus (Carter et al., 2013; Wu et al., 2012a), have been identified in this process. Our data showed that PVH Oxt neurons mediated appetite suppression signals. Previous studies have shown that both oxytocinergic neurites and Oxtr-expressing neurons can be found in various brain and spinal cord regions (Jurek and Neumann, 2018; Lefevre et al., 2021; Newmaster et al., 2020; Oti et al., 2021; Zhang et al., 2021). In the present study, by AAV-Cre-mediated cKO, we found that Oxtr expressed by neurons in the posterior hypothalamic regions, especially those in the ARH, mediates appetite suppression signals (Figures 5 and 6). Our data are, therefore, generally consistent with the view that Oxtr-expressing neurons in the ARH evoke satiety signaling (Fenselau et al., 2017; Maejima et al., 2014); however, we do not exclude the possibility that OxtR in the other parts of the posterior hypothalamus, such as the VMH (Leng et al., 2008; Viskaitis et al., 2017) and medulla (Ong et al., 2017), also contributes to appetite suppression. Collectively, we suggest that one of the output pathways of the PVH for body weight homeostasis is mediated by Oxt signaling-based modulation of other hypothalamic appetite regulation systems.
We also showed that ip administration of Oxt can mitigate the overeating phenotype caused by the PVH Oxt cKO model (Figure 4). Together with the importance of OxtR signaling in the ARH, one possibility is that the primary hypothalamic neurons that are located outside the blood-brain barrier (Yulyaningsih et al., 2017) directly receive ip-injected Oxt and transmit appetite suppression signals. Alternatively, OxtR-expressing neurons in the peripheral nervous system, such as those in the vagal sensory neurons transmitting intentional appetite suppression signals (Bai et al., 2019), may indirectly modify feeding. Future studies should further dissect the responsible cell types and physiological functions of OxtR signaling in the ARH. Recent advances in the real-time imaging of OxtR activities, for example, with a circularly permuted green fluorescent protein binding to OxtR (Ino et al., 2022; Qian et al., 2022), would be useful for delineating the circuit mechanism and spatiotemporal dynamics of the Oxt-mediated suppression of hyperphagic obesity, such as by pinpointing the site of Oxt release.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (mouse, male) | Oxt KO | Inada et al., 2022 | #CDB0204E | |
| Strain, strain background (mouse, male) | Oxt cKO (floxed) | Inada et al., 2022 | #CDB0116E | |
| Strain, strain background (mouse, male) | Oxtrflox/flox | Takayanagi et al., 2005 | ||
| Recombinant DNA reagent | AAV9-hSyn-Cre | Addgene | RRID:Addgene_105555-AAV9 | |
| Recombinant DNA reagent | AAV2-CMV-Cre-GFP | University of North Carolina viral core | https://www.med.unc.edu/genetherapy/vectorcore/in- stock-aav-vectors/reporter-vectors/ | |
| Commercial assay or kit | RNAscope Multiplex Fluorescent Reagent Kit | Advance Cell Diagnostics | 323110 | |
| Commercial assay or kit | RNAscope Mm-OXTR | Advance Cell Diagnostics | 412171 | |
| Software, algorithm | Igor Pro | Wavemetrics | RRID: SCR_000325 | |
| Software, algorithm | ImageJ | NIH | RRID: SCR_003070 |
Animals
All experiments were conducted with virgin male mice. Animals were housed under a 12 hr light/12 hr dark cycle with ad libitum access to water and standard mouse pellets (MFG; Oriental Yeast, Shiga, Japan; 3.57 kcal/g). Wild-type C57BL/6J mice were purchased from Japan SLC (Hamamatsu, Japan). Oxt KO (Accession No. CDB0204E) and cKO (Accession No. CDB0116E) lines (listed at http://www2.clst.riken.jp/arg/mutant%20mice%20list.html) were generated and validated previously (Inada et al., 2022). The Oxtrflox/flox mouse line has been described (Takayanagi et al., 2005). Oxtr KO mice were generated by injecting Cre mRNA into the Oxtrflox/flox zygotes. We used only the mice that had the deletion allele without the flox allele by genotype PCR in the analysis. We confirmed the result of Figure 5—figure supplement 2 in a small number of Oxtr KO mice that had been generated by conventional crossing from Oxtr flox mice. All animal procedures followed the animal care guidelines approved by the Institutional Animal Care and Use Committee of the RIKEN Kobe branch.
We chose the Oxt flox/null model to increase the efficiency of cKO (Figures 1—4). If we had used the Oxt flox/flox mice for cKO, because of high Oxt gene expression levels, a small fraction of the flox alleles that do not experience recombination would easily mask the phenotypes. Because flox/null alone (without Cre) has no phenotype (Figure 1F), we could justify the use of the flox/null model. Regarding Oxtr cKO (Figures 5 and 6), we chose the flox/flox model because the haploinsufficiency gene effect of Oxtr has been reported, at least in the context of social behaviors (Sala et al., 2013).
Stereotactic viral injections
Request a detailed protocolWe obtained the AAV serotype 9 hSyn-Cre from Addgene (#105555; titer: 2.3×1013 genome particles/mL) and the AAV serotype 2 CMV-Cre-GFP from the University of North Carolina viral core (7.1×1012 genome particles/mL). To target the AAV or saline (vehicle) into a specific brain region, stereotactic coordinates were defined for each brain region based on the Allen Mouse Brain Atlas (Lein et al., 2007). Mice were anesthetized with 65 mg/kg ketamine (Daiichi Sankyo, Tokyo, Japan) and 13 mg/kg xylazine (X1251; Sigma-Aldrich) via ip injection and head-fixed to stereotactic equipment (Narishige, Tokyo, Japan). The following coordinates were used (in mm from the bregma for anteroposterior [AP], mediolateral [ML], and dorsoventral [DV]): PVH, AP –0.8, ML 0.2, DV 4.5; SO, AP –0.7, ML 1.2, DV 5.5; LHA, AP –2.0, ML 1.2, DV 5.2; ARH, AP –2.0, ML 0.2, DV 5.8. We defined the anterior and posterior hypothalamus by the following coordinates: anterior, AP +0.2, ML 0.2, DV 5.2; posterior, AP –1.7, ML 1.0, DV 5.2. The injected volume of AAV was 200 nL at a speed of 50 nL/min. After viral injection, the animal was returned to the home cage. In Figure 4, 100 μL of Oxt (1910, Tocris) dissolved in saline (vehicle) at 500 μM or 1 mM was ip-injected once every 3 days from the next day of AAV-Cre injection.
Measurement of food intake
Request a detailed protocolFood intake was measured by placing pre-weighted food pellets on the plate of a cage and reweighing them. In all the experiments, except those in Figure 4—figure supplement 1E–H, daily food intake was measured as follows: 200 g of food pellets were placed and food intake was measured once a week (weekly food intake). Daily food intake was calculated by dividing the weekly food intake by 7 (days) and reported with significance digits of 0.1 g. In Figure 4—figure supplement 1E–H, after 6 hr of fasting, 100 μL of Oxt dissolved in saline (vehicle) at 500 μM or 1 mM was ip-injected. Then, 80.0 g of food was placed and food intake was measured in units of 0.1 g after 1, 3, and 5 hr.
Fluorescent ISH
Request a detailed protocolFluorescent ISH was performed as previously described (Inada et al., 2022; Ishii et al., 2017). In brief, mice were anesthetized with sodium pentobarbital and perfused with PBS followed by 4% PFA in PBS. The brain was post-fixed with 4% PFA overnight. Twenty µm coronal brain sections were made using a cryostat (Leica). The following primer sets were used in this study: Cre forward, CCAAGAAGAAGAGGAAGGTGTC; Cre reverse, ATCCCCAGAAATGCCAGATTAC; Oxt forward, AAGGTCGGTCTGGGCCGGAGA; and Oxt reverse, TAAGCCAAGCAGGCAGCAAGC. Fluoromount (K024; Diagnostic BioSystems) was used as a mounting medium. Brain images were acquired using an Olympus BX53 microscope equipped with a ×10 (NA 0.4) objective lens. Cells were counted manually using the ImageJ Cell Counter plugin. In Figures 1C and 2C, cells were counted by an experimenter who was blind to the experimental conditions.
RNAscope assay
Request a detailed protocolOxtr mRNA was visualized by the RNAscope Multiplex Fluorescent Reagent Kit (323110; Advance Cell Diagnostics [ACD]) according to the manufacturer’s instructions. In brief, 20 µm coronal brain sections were made using a cryostat (Leica). A probe against Oxtr (Mm-OXTR, 412171, ACD) was hybridized in a HybEZ Oven (ACD) for 2 hr at 40°C. Then, the sections were treated with TSA-plus Cyanine 3 (NEL744001KT; Akoya Biosciences; 1:1500). Fluoromount (K024; Diagnostic BioSystems) was used as a mounting medium. Images subjected to the analysis were acquired using an Olympus BX53 microscope equipped with a ×10 (NA 0.40) or ×20 (NA 0.75) objective lens, as shown in Figures 5E, 6E and F. Figure 5F was obtained with a confocal microscope (LSM780, Zeiss) equipped with a ×63 oil-immersion objective lens (NA 1.40). RNAscope dots were counted manually using the ImageJ Cell Counter plugin.
Measurements of the weight of livers and stomachs
Request a detailed protocolMice were anesthetized with sodium pentobarbital. Livers and stomachs were obtained without perfusion and their weight was immediately measured. Before measurement, the stomach was gently pressed to eject the remaining contents.
Plasma measurements
Request a detailed protocolMice were fed ad libitum before blood sampling. Mice were anesthetized with isoflurane and blood was collected from the heart. EDTA was used to prevent blood coagulation. Plasma concentrations of glucose, triglycerides, and leptin were measured by the enzymatic method, HK-G6PDH, and ELISA, respectively, through a service provided by Oriental Yeast (Shiga, Japan).
Data analysis
Request a detailed protocolAll mean values are reported as the mean ± SEM. The statistical details of each experiment, including the statistical tests used, the exact value of n, and what n represents, are shown in each figure legend. The p-values are shown in each figure legend or panel; nonsignificant values are not noted. In Figures 1G and 2E, exponential fit was calculated by Igor (WaveMetrics).
Data availability
All data generated or analyzed during this study are included in the manuscript and the figure supplement.
References
-
Evidence that paraventricular nucleus oxytocin neurons link hypothalamic leptin action to caudal brain stem nuclei controlling meal sizeAmerican Journal of Physiology. Regulatory, Integrative and Comparative Physiology 287:R87–R96.https://doi.org/10.1152/ajpregu.00604.2003
-
Oxytocin receptor gene polymorphism and low serum oxytocin level are associated with hyperphagia and obesity in adolescentsInternational Journal of Obesity 45:2064–2073.https://doi.org/10.1038/s41366-021-00876-5
-
A neural basis for melanocortin-4 receptor-regulated appetiteNature Neuroscience 18:863–871.https://doi.org/10.1038/nn.4011
-
The oxytocin receptor: from intracellular signaling to behaviorPhysiological Reviews 98:1805–1908.https://doi.org/10.1152/physrev.00031.2017
-
Oxytocinergic feedback circuitries: an anatomical basis for neuromodulation of social behaviorsFrontiers in Neural Circuits 15:688234.https://doi.org/10.3389/fncir.2021.688234
-
Oxytocin and appetiteProgress in Brain Research 170:137–151.https://doi.org/10.1016/S0079-6123(08)00413-5
-
Oxytocin-the sweet hormone?Trends in Endocrinology and Metabolism 28:365–376.https://doi.org/10.1016/j.tem.2017.02.007
-
The anorexigenic neural pathways of oxytocin and their clinical implicationNeuroendocrinology 107:91–104.https://doi.org/10.1159/000489263
-
A distributed network for social cognition enriched for oxytocin receptorsThe Journal of Neuroscience 36:2517–2535.https://doi.org/10.1523/JNEUROSCI.2409-15.2016
-
Role of oxytocin in the control of stress and food intakeJournal of Neuroendocrinology 31:e12700.https://doi.org/10.1111/jne.12700
-
Amygdalohippocampal area neurons that project to the preoptic area mediate infant-directed attack in male miceThe Journal of Neuroscience 40:3981–3994.https://doi.org/10.1523/JNEUROSCI.0438-19.2020
-
Effect of paraventricular nucleus lesions on body weight, food intake and insulin levelsBehavioural Brain Research 22:265–281.https://doi.org/10.1016/0166-4328(86)90071-9
-
Three pillars for the neural control of appetiteAnnual Review of Physiology 79:401–423.https://doi.org/10.1146/annurev-physiol-021115-104948
-
The role of PVH circuits in leptin action and energy balanceAnnual Review of Physiology 78:207–221.https://doi.org/10.1146/annurev-physiol-021115-105347
-
Oxytocin’s inhibitory effect on food intake is stronger in obese than normal-weight menInternational Journal of Obesity 40:1707–1714.https://doi.org/10.1038/ijo.2016.149
-
RNAscope: a novel in situ RNA analysis platform for formalin-fixed, paraffin-embedded tissuesThe Journal of Molecular Diagnostics 14:22–29.https://doi.org/10.1016/j.jmoldx.2011.08.002
-
Do oxytocin neurones affect feeding?Journal of Neuroendocrinology 33:e13035.https://doi.org/10.1111/jne.13035
Article and author information
Author details
Kazunari Miyamichi

Funding
Japan Society for the Promotion of Science (KAKENHI 19J00403)
- Kengo Inada
Japan Society for the Promotion of Science (KAKENHI 19K16303)
- Kengo Inada
Japan Science and Technology Agency (CREST JPMJCR2021)
- Kazunari Miyamichi
Japan Society for the Promotion of Science (KAKENHI 20K20589)
- Kazunari Miyamichi
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We wish to thank Mitsue Hagihara for the extensive technical support, the Laboratory for Comprehensive Bioimaging for the microscopy services, and the members of the Miyamichi lab for their comments on an earlier version of this manuscript. This work was supported by the RIKEN Special Postdoctoral Researchers Program, a grant from the Kao Foundation for Arts and Sciences, and JSPS KAKENHI (19J00403 and 19K16303) to KI, and the JST CREST program (JPMJCR2021) and JSPS KAKENHI (20K20589) to KM.
Ethics
All animal procedures followed animal care guidelines approved by the Institutional Animal Care and Use Committee of the RIKEN Kobe branch (#A2017-15-12).
Copyright
© 2022, Inada et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,349
- views
-
- 381
- downloads
-
- 37
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 37
- citations for umbrella DOI https://doi.org/10.7554/eLife.75718
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Oxytocin signaling in the posterior hypothalamus prevents hyperphagic obesity in mice
eLife 11:e75718.
https://doi.org/10.7554/eLife.75718




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}