Cystatin F (Cst7) drives sex-dependent changes in microglia in an amyloid-driven model of Alzheimer’s disease
- UK Dementia Research Insitute at The University of Edinburgh, United Kingdom
- Centre for Inflammation Research, Institute for Regeneration and Repair, The University of Edinburgh, United Kingdom
- Simons Initiative for the Developing Brain, University of Edinburgh, United Kingdom
- Department of Neurology and Neurological Science, Graduate School of Medicine, Tokyo Medical and Dental University, Japan
- Laboratory for Proteolytic Neuroscience, RIKEN Brain Science Institute, Japan
- Department of Neuroscience and Pathobiology, Research Institute of Environmental Medicine, Nagoya University, Japan
Abstract
Microglial endolysosomal (dys)function is strongly implicated in neurodegenerative disease. Transcriptomic studies show that a microglial state characterised by a set of genes involved in endolysosomal function is induced in both mouse Alzheimer’s disease (AD) models and human AD brain, and that the emergence of this state is emphasised in females. Cst7 (encoding cystatin F) is among the most highly upregulated genes in these microglia. However, despite such striking and robust upregulation, the function of Cst7 in neurodegenerative disease is not understood. Here, we crossed Cst7-/- mice with the AppNL-G-F mouse to test the role of Cst7 in a model of amyloid-driven AD. Surprisingly, we found that Cst7 plays a sexually dimorphic role regulating microglia in this model. In females, Cst7-/-AppNL-G-F microglia had greater endolysosomal gene expression, lysosomal burden, and amyloid beta (Aβ) burden in vivo and were more phagocytic in vitro. However, in males, Cst7-/-AppNL-G-F microglia were less inflammatory and had a reduction in lysosomal burden but had no change in Aβ burden. Overall, our study reveals functional roles for one of the most commonly upregulated genes in microglia across disease models, and the sex-specific profiles of Cst7-/--altered microglial disease phenotypes. More broadly, the findings raise important implications for AD including crucial questions on sexual dimorphism in neurodegenerative disease and the interplay between endolysosomal and inflammatory pathways in AD pathology.
Editor's evaluation
This study presents a valuable finding on the function of the gene Cst7 in sex-divergent pathological changes in microglia in a mouse model of amyloid-driven Alzheimer's disease. The evidence supporting the claims of the authors is solid, although the study would be further strengthened by validation of some of the key differentially expressed genes identified in RNA-sequencing experiments. Overall, this study offers new insight into the functional role of CST7 that is upregulated in a subset of disease-associated microglia in AD models and the human brain that will be of interest to neuroimmunologists and neuroscientists working on microglia in health and disease.
https://doi.org/10.7554/eLife.85279.sa0Introduction
Alzheimer’s disease (AD) is a chronic neurodegenerative disease characterised pathologically by build-up of protein aggregates such as amyloid beta (Aβ) and hyperphosphorylated tau. AD is also characterised by reactive glial cells, including the emergence of altered microglia. Microglia are the predominant immune cells of the brain parenchyma and play numerous roles in development and homeostasis (Colonna and Butovsky, 2017). In recent years, microglia have become heavily implicated in the pathogenesis of AD, in part due to accumulating genetic and epigenetic evidence that single nucleotide polymorphisms leading to altered risk for late-onset AD (LOAD) are enriched in microglial-expressed genes associated with endolysosomal function and lipid handling (Podleśny-Drabiniok et al., 2020). This implies that microglia may play a causal role in the disease and supports the theory that microglia may be targeted to treat AD. In addition to genetics, sex is a major risk factor for AD, with women up to twice as likely to be diagnosed than men (Podcasy and Epperson, 2016). This sex bias is not entirely due to increased longevity in females, as increased risk remains after adjusting for lifespan, and may be due to interaction effects between sex/gender and genetics (Gamache et al., 2020). The mechanisms for this, and possible interplay between microglia and sex are poorly understood.
Recently, an altered state of microglia has been identified in AD models that arises dependent on AD risk gene TREM2. These cells, termed disease-associated microglia (DAM)/microglial neurodegenerative phenotype (MGnD)/activated response microglia (ARM) (Keren-Shaul et al., 2017; Krasemann et al., 2017; Sala Frigerio et al., 2019), are characterised by downregulation of microglial ‘homeostatic’ genes Tmem119 and P2ry12 and upregulation of genes such as Apoe, Spp1, Itgax, Clec7a, and Cst7. Notably, the emergence of this subset may be accelerated in female mice compared to male (Sala Frigerio et al., 2019). DAM/MGnD/ARM are believed to be highly phagocytic cells due to the enrichment of phagocytic and lipid-handling pathways in their gene sets (Deczkowska et al., 2018). However, the precise functions of many of the genes that make up DAM/MGnD/ARM are poorly understood.
Cst7, coding for the protein cystatin F (CF), is amongst the most robustly upregulated genes in the DAM/MGnD/ARM signature. Cst7/CF expression appears to be driven by amyloid plaques as upregulation at both the RNA and protein level is spatially localised to plaques (Ofengeim et al., 2017; Chen et al., 2020). CF is atypical in that it is localised to the endolysosomal compartment where it acts as an endogenous inhibitor of cysteine proteases such as cathepsin L and C (Hamilton et al., 2008). However, little is understood about the mechanistic role of CF in the context of central nervous system (CNS) disease. Some studies have suggested CF may play a protective role in a demyelination model by inhibiting cathepsin C (Liang et al., 2016) or negatively regulating microglial phagocytosis (Kang et al., 2018; Popescu et al., 2023) but the possible disease-modifying role of Cst7/CF in amyloid-driven neurodegenerative disease is unknown. Additionally, although there is some evidence that Cst7 itself is upregulated to a greater extent in female vs. male mice (Kang et al., 2018; Sala Frigerio et al., 2019; Guillot-Sestier et al., 2021), the sex-specific role of Cst7 in disease has not been investigated.
Here, we set out to investigate what, if any, disease-modifying role Cst7/CF may play in an amyloid-driven AD mouse model in both females and males. We first compiled data from published datasets to show that Cst7 is indeed robustly and consistently upregulated in microglia across numerous amyloid-driven models of AD. We then deleted Cst7 in male and female AppNL-G-F knock-in mice which showed that Cst7 drives sex-dependent changes in microglia at transcript, protein, and functional level. Cst7 deletion in male mice led to downregulation of inflammatory transcripts and reduced lysosomal burden but had no effect on microglial Aβ burden in vivo or plaque deposition. However, in females, Cst7 deletion led to increased expression of endolysosomal genes, increased lysosomal burden, and increased microglial Aβ burden in vivo. We further investigated this in vitro and showed that this is due to increased phagocytosis rather than impaired degradation. These data bring important insight to the functional role of Cst7, a key hallmark gene within the DAM/MGnD/ARM disease microglia signature. They also highlight that pathology-influencing effects are sex-dependent, underlining the interaction between sex and microglial function relevant to neurodegenerative disease.
Results
Cst7/CF is upregulated in female and male microglia in murine models of AD
Cst7 is often detected in RNASeq or scRNASeq databases as significantly upregulated in disease models. However, an integrated analysis of Cst7 expression in disease models stratified by sex is lacking. Therefore, we searched publicly available datasets for Cst7 expression in microglia in mouse models of AD. We integrated data from six studies in which microglial Cst7 expression was profiled (Figure 1A). Expression data were restricted to microglia by use of techniques such as fluorescence-activated cell sorting (FACS) (Orre et al., 2014; Wang et al., 2015; Srinivasan et al., 2020), immunomagnetic separation (Guillot-Sestier et al., 2021), RiboTag translational profiling (Kang et al., 2018), or single-cell RNA sequencing (Sala Frigerio et al., 2019). We were able to collect data on Cst7 expression in males vs. females in the APP/PS1 (Guillot-Sestier et al., 2021) and AppNL-G-F model (Sala Frigerio et al., 2019). In all models, Cst7 was dramatically upregulated in microglia in amyloid-driven disease and in models where we were able to compare sexes, Cst7 appeared marginally more highly expressed in female disease brains than in male. This is consistent with literature suggesting the DAM/ARM/MGnD microglial subset is accelerated in female mice (Sala Frigerio et al., 2019). We further investigated sexual dimorphism in microglial states by sub-setting ARMs from Sala-Frigerio et al. and discovered, alongside sex-chromosome genes, endolysosomal genes such as Spp1 and Gpnmb amongst Cst7 as marginally more expressed in female ARMs vs. male ARMs (Figure 1—figure supplement 1A). To investigate cell-specific expression of Cst7, we mined sequencing databases and found Cst7 is expressed almost exclusively in disease-derived microglial clusters with some minimal expression in homeostatic microglia and negligible expression in other CNS or immune cell subtypes (Zhang et al., 2014; Van Hove et al., 2019; Zeng et al., 2023; Figure 1—figure supplement 1B–D). Finally, we investigated the expression of Cst7 in intact tissue by conducting in situ hybridisation for Cst7 with markers for IBA1 and Aβ plaques for localisation. We found that Cst7 was dramatically upregulated in disease where expression decorated plaques in AppNL-G-F brains (Figure 1B). Furthermore, expression was significantly enriched around plaques (Figure 1C and D) and almost exclusively localised to IBA1+ cells surrounding plaques (Figure 1E and F). Together, these data validate Cst7 as a key marker gene of disease-induced reactive microglia, notably of the DAM/MGnD/ARM state, across cerebral amyloidosis models in both sexes, and suggest that its disease-induced upregulation is related temporally and spatially to Aβ plaque pathology.
Figure 1 with 1 supplement see all
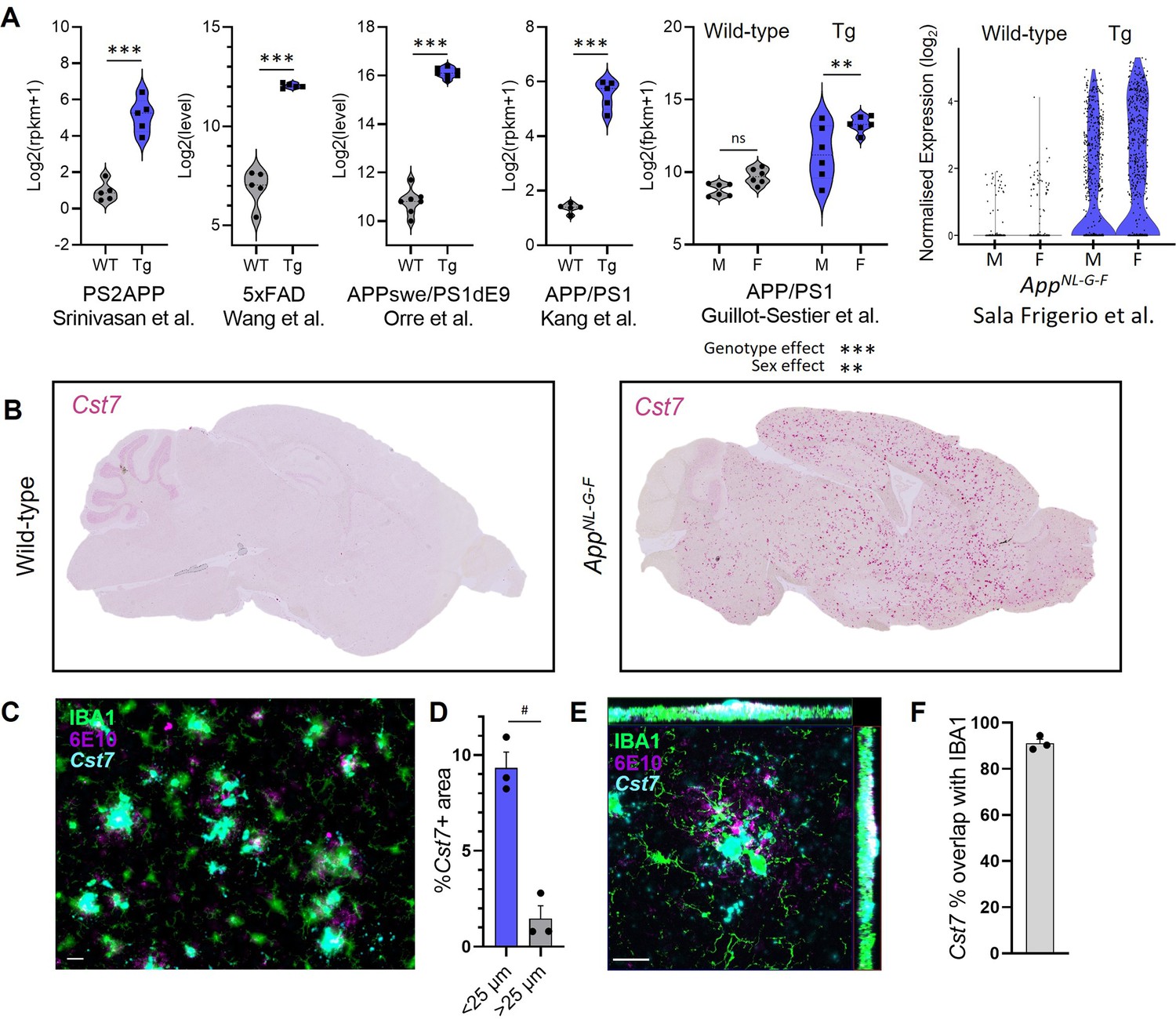
Cst7/cystatin F is upregulated in microglia in murine models of Alzheimer’s disease (AD).
(A) Analysis of publicly available microglial RNASeq databases for Cst7 expression in amyloid-driven models. Background-matched control (grey) or relevant disease model (blue). (B) Brightfield slide-scanned images of wild-type (left) and AppNL-G-F (right) sagittal brain sections stained by in situ hybridisation for Cst7 (red). (C) Example image from Cst7 fluorescence in situ hybridisation (FISH) experiment stained with IBA1 (green), 6E10 (magenta), and Cst7 (cyan). (D) Quantification of (C) showing % Cst7+ area in areas within 25 µm (blue) of 6E10+ plaques or outside (grey). (E) Hi-mag confocal microscopy maximum intensity projection showing co-localisation of Cst7 with IBA1 around plaques. (F) Quantification of (E) showing % Cst7 overlap with IBA1. n=3–7. **p<0.01, ***p<0.001 by unpaired t-test or two-way ANOVA with Tukey’s multiple comparisons post hoc test. #p<0.05 by paired t-test. Scale bars are 20 µm.
-
Figure 1—source data 1
Source data associated with Figure 1.
(D) Quantification of (C) showing % Cst7+ area in areas within 25 µm of 6E10+ plaques or outside. (F) Quantification of (E) showing % Cst7 overlap with IBA1.
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig1-data1-v1.xlsx
Cst7 deletion leads to sex-dependent transcriptomic changes in microglia in the AppNL-G-F model of amyloid-driven AD
As Cst7 is so robustly upregulated in microglia in amyloid-driven disease and some studies suggest this to be greater in female mice, we next aimed to investigate whether Cst7/CF plays a role in regulating microglia in disease and whether this regulation could be sex-dependent. Therefore, we crossed Cst7-/- mice with the AppNL-G-F model of amyloid-driven AD (Saito et al., 2014) to generate background-matched non-disease (AppWt/WtCst7+/+), disease (AppNL-G-FCst7+/+) and disease knockout (AppNL-G-FCst7-/-) mice. As Cst7 expression is limited to the myeloid compartment in the brain (Zhang et al., 2014), this study primarily tests the role of microglial/border-associated macrophages in amyloid-driven disease. We first investigated the role of Cst7/CF in regulating the microglial response in disease by unbiased analysis of the microglial transcriptome. Mice were aged to 12 months to drive Aβ plaque burden and a DAM/ARM microglial profile that includes marked upregulation of Cst7 before microglia were isolated from hemi-brains by FACS (Figure 2A). Microglia were defined by FSC/SSC profile, singlet, live, CD11b+, Ly6C-, CD45+ (Figure 2—figure supplement 1A) and sorted along with phenotypic markers P2Y12, MHCII, and CD11c (Figure 2—figure supplement 1) before RNA purification, sequencing, and analysis using DESeq2 (Love et al., 2014).
Figure 2 with 3 supplements see all
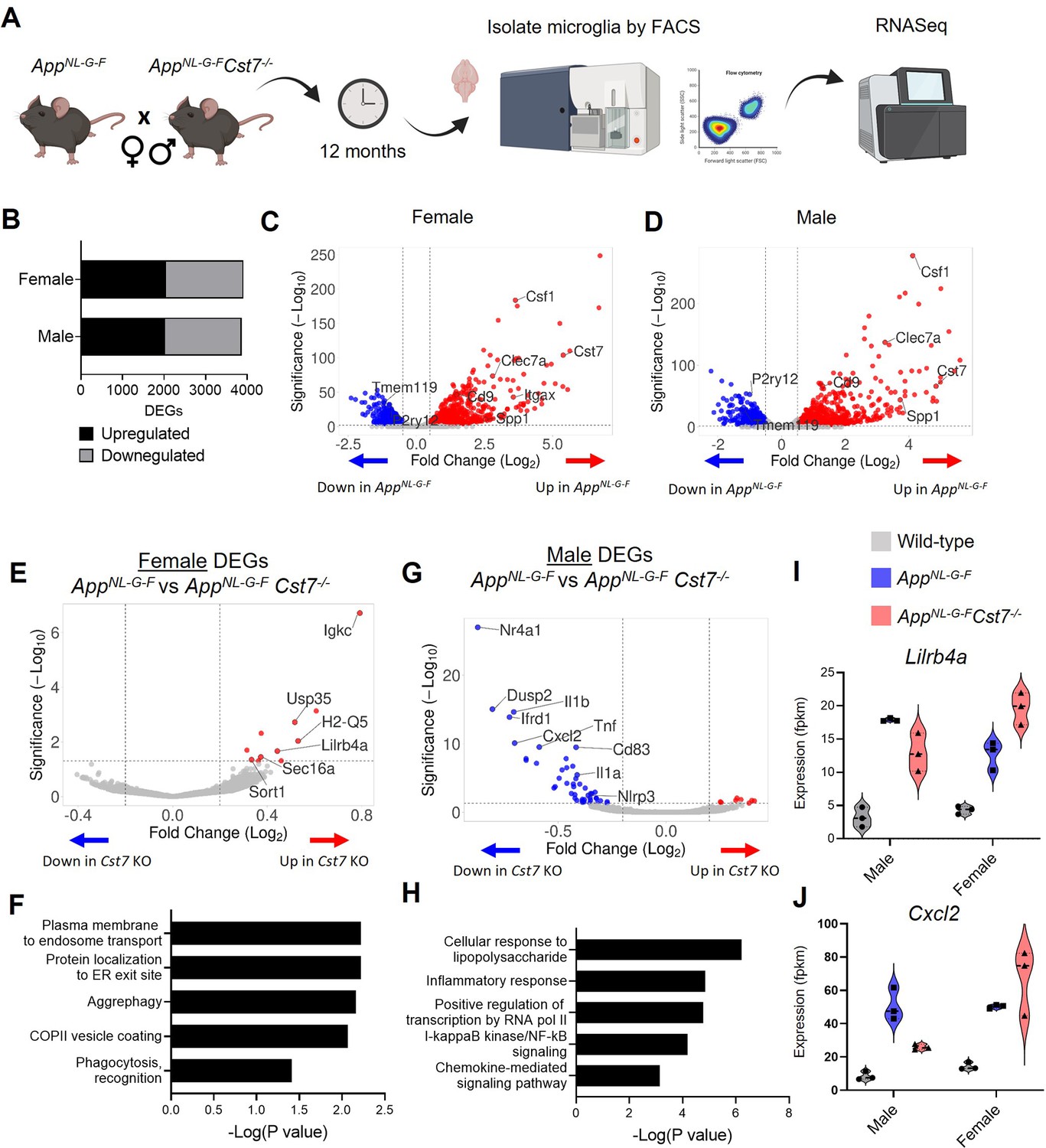
Cst7 deletion leads to sex-dependent transcriptomic changes in microglia in the AppNL-G-F model of amyloid-driven Alzheimer’s disease (AD).
(A) Study design schematic. (B) Differentially expressed genes (DEGs) between wild-type and AppNL-G-F microglia in male and female mice. (C) Volcano plot of RNASeq from wild-type vs. AppNL-G-F female microglia. Points in red are significantly upregulated in disease. Points in blue are significantly downregulated. Selected genes are annotated. (D) Volcano plot of RNASeq from wild-type vs. AppNL-G-FCst7+/+ male microglia. (E) Volcano plot of RNASeq from female AppNL-G-FCst7+/+ vs. AppNL-G-FCst7-/- microglia. Points in red are significantly upregulated in AppNL-G-FCst7-/-. Points in blue are significantly downregulated. Selected genes are annotated. (F) Selected GO:BP (Gene Ontology:biological process) terms that are significantly enriched in the upregulated genes from (E) and corresponding significance p-value. (G) Volcano plot of RNASeq from male AppNL-G-FCst7+/+ vs. AppNL-G-FCst7-/- microglia. (H) Selected GO:BP terms that are significantly enriched in the upregulated genes from (G) and corresponding significance p-value. (I and J) Example gene expression (fpkm) from RNASeq of microglia isolated from male and female wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice. Selected genes are Lilrb4 (I) and Cxcl2 (J).
-
Figure 2—source data 1
Source data associated with Figure 2.
(I and J) Example gene expression (fpkm) from RNASeq of microglia isolated from male and female wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice. Selected genes are Lilrb4 (I) and Cxcl2 (J).
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig2-data1-v1.xlsx
-
Figure 2—source data 2
Differentially expressed genes (DEGs) between male and female AppNL-G-F and AppNL-G-FCst7-/- mice (selected genes in bold).
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig2-data2-v1.docx
We first assessed disease-induced microglial changes by comparing AppWt/WtCst7+/+ and AppNL-G-FCst7+/+ microglia in male and female mice. AppNL-G-F knock-in led to numerous changes in gene transcription with 2022 genes upregulated and 1849 downregulated in males and 2053 upregulated and 1857 downregulated in females (Figure 2B). Differentially regulated genes were mostly shared between males and females but we did observe uniquely regulated genes between sexes (Figure 2—figure supplement 2B). As expected, upregulated genes included DAM/ARM genes such as Clec7a, Spp1, Itgax, and Csf1, whilst downregulated genes included microglial ‘homeostatic’ genes P2ry12, Tmem119, and Gpr34 in both sexes (Figure 2C and D). Cst7 was among the top 10 most highly upregulated genes in males and females. Interestingly, although there were relatively few differentially expressed genes (DEGs) between male and female mice in the AppNL-G-FCst7+/+ group (Figure 2—source data 2), these genes did include DAM/ARM genes such as Gpnmb, Spp1, and Ctse. This is consistent with the observation that the DAM/ARM profile is accelerated in females compared to males (Figure 1A; Sala Frigerio et al., 2019).
Having confirmed the upregulation of the Cst7 and the DAM/ARM profile in this model, we next tested the role of Cst7 in disease by comparing differential expression between the AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- groups in both sexes. Importantly, Cst7 itself was the most DEG in both sexes, confirming successful deletion (Figure 2—figure supplement 2A). Cst7 deletion led to significant changes in microglial transcriptome. In females, Cst7 knockout led to upregulation of 15 genes and downregulation of 2 genes (including Cst7) (Figure 2E). Despite the modest number of DEGs, we observed that many of the upregulated genes appeared to be involved in endolysosomal-related processes such as protein trafficking to organelles (Sort1, Sec16a) and antigen presentation (Lilrb4a, Igkc). Enrichment analyses revealed that female AppNL-G-FCst7-/- microglia were significantly enriched in GO:BP (Gene Ontology: biological process) terms such as ‘plasma membrane to endosome transport’, ‘aggrephagy’, and ‘phagocytosis’ against their AppNL-G-FCst7+/+ counterparts (Figure 2F).
Next, we tested whether Cst7 knockout led to the same effect in males as in females. Surprisingly, we observed that male AppNL-G-FCst7-/- microglia exhibited markedly different transcriptome compared to AppNL-G-F with upregulation of 8 genes and downregulation of 51 genes (including Cst7) (Figure 2G). In contrast to females, downregulated genes in males were dominated by classical proinflammatory mediators such as Il1b, Il1a, Tnf, and Cxcl2 and enrichment analyses revealed downregulation of GO:BP terms such as ‘cellular response to lipopolysaccharide’, inflammatory response’, and ‘NF-kappaB signaling’ (Figure 2H). There was very little overlap between Cst7-regulated genes in males and females with only two genes shared including Cst7 (Figure 2—figure supplement 2C). Notably, many disease-induced genes followed an intriguing pattern of being suppressed in AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+ in males but potentiated in AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+ in females. Examples of this are immunoglobulin receptor gene Lilrb4a and inflammatory gene Cxcl2 (Figure 2I and J). Interestingly, Cst7 also appeared to drive a sex-genotype interaction effect. We determined male vs. female DEGs in AppNL-G-FCst7+/+ and their AppNL-G-FCst7-/- counterparts. In AppNL-G-FCst7+/+ microglia, there were 33 DEGs between male vs. female approximately evenly split between transcripts enriched in males vs. those enriched in females, whereas in AppNL-G-FCst7-/- we observed 240 DEGs dominated by those enriched in females (Figure 2—figure supplement 2D). These transcripts included inflammatory genes such as Il1b, Tnf, and Cxcl2.
Finally, we sought to validate our findings through complimentary methods and confirm the gene changes we observe are microglia-expressed. We used fluorescence in situ hybridisation (FISH) coupled with immunostaining to measure expression of Lilrb4a in non-processed brain tissue. We confirmed expression of Lilrb4a in microglia specifically around plaques and validated our RNASeq data showing an identical expression pattern to that observed previously with increased expression of microglial Lilrb4a in AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+ in females but not in males (Figure 2—figure supplement 3E and F). We also assessed expression of key hit genes Lilrb4a and Il1b by performing qPCR in microglia isolated from non-diseased wild-type vs Cst7-/- mice to confirm disease-specific regulation. Indeed, we did not see the same pattern as described in RNASeq with upregulation in disease and further upregulation in Cst7-/- in females (Lilrb4a) or downregulation in Cst7-/- males (Il1b) (Figure 2—figure supplement 3A). In fact, there were no significant differences between groups (Figure 2—figure supplement 3B).
Together, these data show that Cst7 drives sex-dependent effects on microglial transcriptome in an amyloid-driven mouse model of AD. Cst7 knockout led to a disease-dependent upregulation of a small set of endolysosomal-enriched genes in female microglia but a striking downregulation of inflammatory genes in male microglia. The number of DEGs between AppNL-G-F males and females was markedly greater in the Cst7-/- state, implying a sex-genotype interaction effect, that is, the impact of Cst7 deficiency on microglial phenotype in AppNL-G-F-driven disease differs quantitatively and qualitatively (e.g. in endolysosomal and inflammatory pathways) according to sex.
Cst7 deletion leads to region and sex-dependent changes in lysosomes in AppNL-G-F mice
Following our investigation into the Cst7-dependent effects on microglia at the transcriptome level, we next sought to study changes at the protein level with spatial resolution. This allowed us to investigate brain region-dependent effects. We took sagittal brain sections from 12-month male and female AppWt/WtCst7+/+, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice and immunostained for IBA1 to visualise microglia/macrophages, anti-Aβ clone 6E10 to visualise Aβ, and, as both previous literature and our RNASeq data in Figure 2 implicated lysosomes in Cst7 function, LAMP2 to visualise lysosomes. We assessed three brain regions: cortex, whole hippocampus, and the dorsal subiculum (Figure 3—figure supplement 1A), to investigate the effects of disease and Cst7 knockout on microglial burden and lysosomal activation. As expected, knock-in of the AppNL-G-F construct led to marked build-up of 6E10+ Aβ plaques, an increase in IBA1+ microglial burden around these plaques, and upregulation of LAMP2 in microglia predominantly in plaque-associated microglia in both males and females (Figure 3A). LAMP2 predominantly co-localised with IBA1, suggesting that expression is predominant in mononuclear phagocytes rather than neuronal cells or macroglia. Next, we compared AppNL-G-FCst7+/+ vs. AppNL-G-FCst7-/- male and female mice. We observed a trend towards increased IBA1+ microglial burden in AppNL-G-FCst7-/- mice in all brain regions, statistically significant in the subiculum (Figure 3B–D), more evidently in male mice. Importantly, this was not only due to an increase in Aβ plaque load (which correlated with microglial burden) as images were quantified in areas of even plaque coverage (Figure 3—figure supplement 1B–D).
Figure 3 with 1 supplement see all
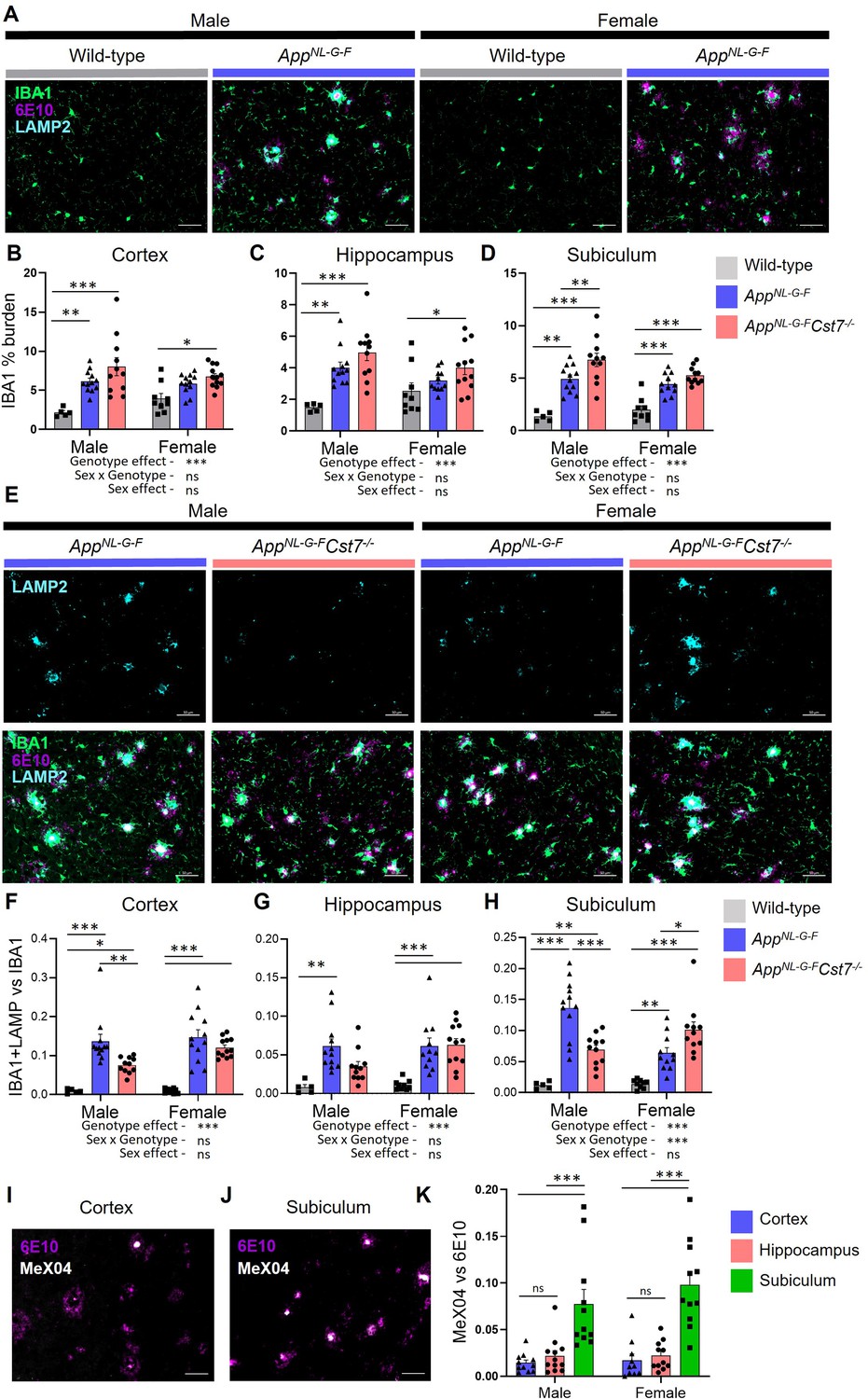
Cst7 deletion leads to region and sex-dependent changes in lysosomes in AppNL-G-F mice.
(A) Example images from male and female wild-type and AppNL-G-F cortex stained with IBA1 (green), 6E10 (magenta), and LAMP2 (cyan). (B–D) IBA1 % coverage from images taken of wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- brains in cortex (B), hippocampus (C), and dorsal subiculum (D). (E) Example images from AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- subicula in male and female mice. Top panel shows LAMP2 (cyan) and bottom panel shows IBA1 (green), 6E10 (magenta), and LAMP2 (cyan) merge. Scale bars are 50 μm. (F–H) Ratio of IBA1/LAMP2 double positive staining vs. IBA1 total staining in the cortex (F), hippocampus (G), and dorsal subiculum (H) of wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice. (I and J) Example images from the cortex (I) and subiculum (J) of an AppNL-G-F mouse stained with 6E10 (magenta) and MeX04 (white). (K) Ratio of MeX04 total staining vs. 6E10 total staining in cortex (blue), hippocampus (red), and subiculum (green) of male and female AppNL-G-F mice. Scale bars are 50 μm. Data points are average from 2 fields of view/mouse (1 for subiculum) and bars are plotted as mean + SEM. n=5–12. *p<0.05, **p<0.01, ***p<0.001 by two-way ANOVA with Tukey’s multiple comparisons post hoc test.
-
Figure 3—source data 1
Source data associated with Figure 3.
(B–D) IBA1 % coverage from images taken of wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- brains in cortex (B), hippocampus (C), and dorsal subiculum (D). (F–H) Ratio of IBA1/LAMP2 double positive staining vs. IBA1 total staining in the cortex (F), hippocampus (G), and dorsal subiculum (H) of wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice. (K) Ratio of MeX04 total staining vs. 6E10 total staining in cortex (blue), hippocampus (red), and subiculum (green) of male and female AppNL-G-F mice.
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig3-data1-v1.xlsx
To investigate the effect of Cst7 deletion on lysosomal activity, we quantified LAMP2 immunostaining and normalised to IBA1 as these markers are highly correlated (Figure 3—figure supplement 1E) and we wanted to remove the confounding factor of a greater IBA1 burden in Cst7 knockout mice (Figure 3B–D). Strikingly, we found in the subiculum that microglial-normalised LAMP2 staining followed the same pattern as many of the genes identified by microglia RNASeq with reduced expression in AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+ males but increased expression in AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+ females (Figure 3E, quantification in Figure 3H). A similar pattern was observed in the cortex and hippocampus but without a significant change in females (Figure 3F and G). We hypothesised that the reason for this region-dependent effect may be due to differences in plaque structure. Aβ plaques typically comprise a β-sheet-containing dense core and a more diffuse outer ring (DeTure and Dickson, 2019). We utilised Congo Red derivative methoxy-X04 (MeX04), which binds selectively to fibrillar β-sheet-rich structures such as the dense cores of amyloid plaques (DeTure and Dickson, 2019), compared to anti-Aβ antibody 6E10, which will bind both dense cores and diffuse outers to study plaque composition in different brain regions of AppNL-G-F mice (Figure 3—figure supplement 1A). MeX04 was injected intraperitoneally 2.5 hr before mice were sacrificed and brains taken for histology. We found that Aβ plaques had a far greater MeX04:6E10 ratio in the subiculum compared to other brain regions (Figure 3I and J, quantified in Figure 3K), most likely caused by earlier Aβ accumulation in this region (Rönnbäck et al., 2011; Gail Canter et al., 2019) that may be dependent on a subpopulation of microglia (Spangenberg et al., 2019). This implies that the sex-dependent effects of Cst7 deletion on microglial lysosomes may be driven by plaque composition/structure, with regions dominated by dense core plaques (indicating further ‘disease progression’) showing greater effect of Cst7 deletion.
Cst7 regulates microglial Aβ burden in female AppNL-G-F mice
After discovering that Cst7 deletion leads to sex-dependent changes in microglia at both the transcriptome and protein level indicative of an altered endolysosomal system, we next further investigated potential functional changes in these altered microglia. A major function of microglia in the context of AD is uptake of Aβ into lysosomes (Grubman et al., 2021). To study this, we utilised MeX04 injection as described previously to label Aβ-containing microglia in the brain. First, we validated that intraperitoneal injection of MeX04 2.5 hr before microglial isolation by FACS can detect Aβ-containing microglia. Wild-type and AppNL-G-F mice were aged to 12 months and injected with either MeX04 or vehicle before microglial isolation by FACS as previously but with additional detection of MeX04 in the DAPI channel. As expected, wild-type+MeX04 and AppNL-G-F+vehicle mice displayed a background MeX04 signal that was shifted only in the AppNL-G-F+MeX04 group (Figure 4—figure supplement 1).
After validating our approach, we next investigated the role of Cst7 on microglial Aβ burden (Figure 4A). Consistent with our RNASeq and histology data suggesting increased lysosomal capacity specifically in female AppNL-G-F mice following deletion of Cst7, we observed a greater Aβ burden in female AppNL-G-FCst7-/- microglia compared to AppNL-G-FCst7+/+ counterparts measured by both % Aβ-positive microglia (Figure 4B) and mean fluorescence intensity of MeX04 on microglia (Figure 4C). Interestingly, despite a reduced expression of lysosomal membrane marker LAMP2 in male AppNL-G-FCst7-/- mice compared to AppNL-G-FCst7+/+ counterparts (Figure 3C and D), we observed no difference in microglial Aβ burden (Figure 4B). This may be due to the counteracting effect of a reduction of inflammatory genes such as Il1b and Nlrp3 (Figure 2G) which have been shown to impair phagocytosis (Heneka et al., 2013; Tejera et al., 2019).
Figure 4 with 1 supplement see all
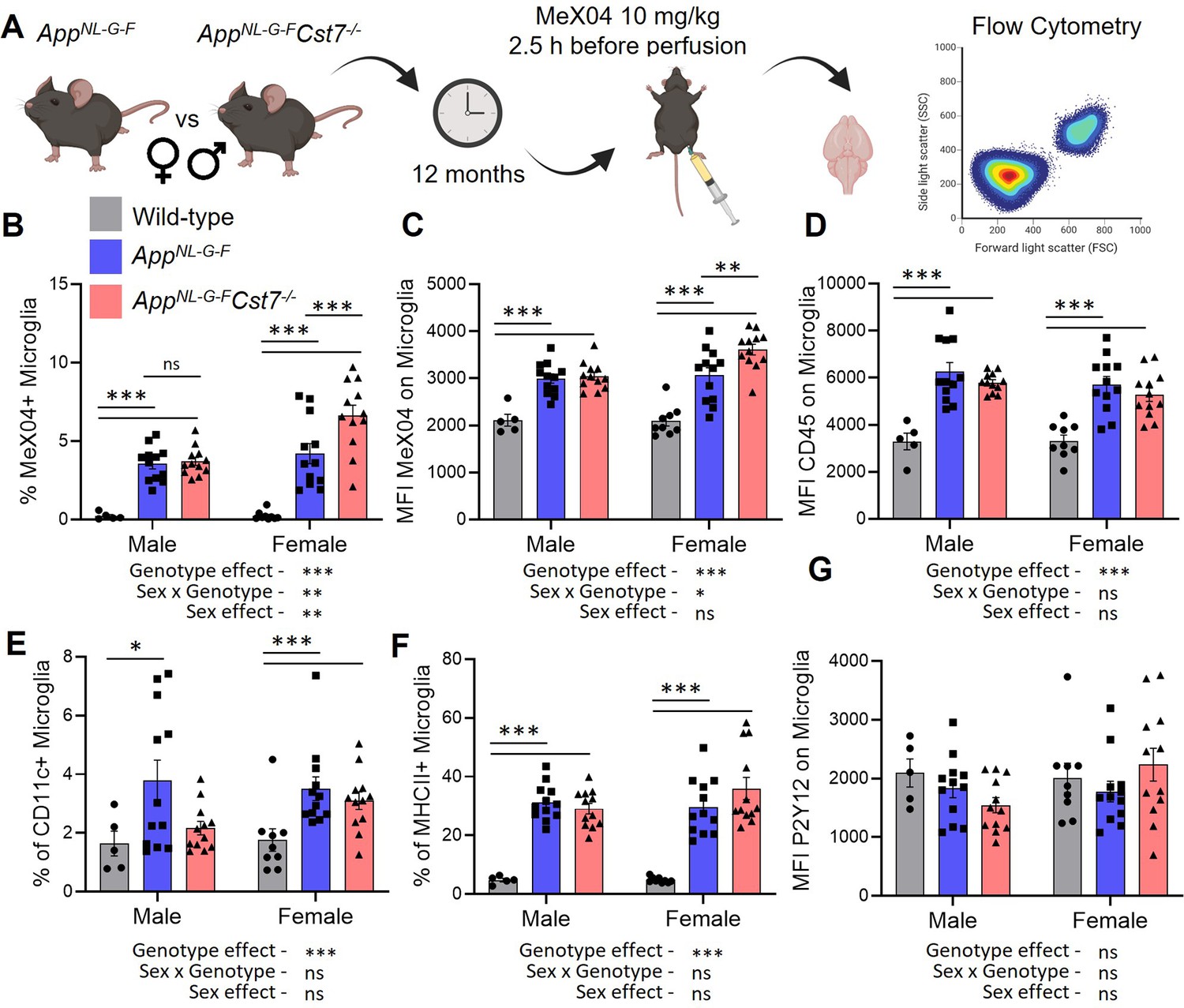
Cst7 regulates microglial Aβ burden in female AppNL-G-F mice.
(A) Study design schematic. (B) % MeX04+ microglia of total microglia in male and female wild-type (grey), AppNL-G-FCst7+/+ (blue), and AppNL-G-FCst7-/- (red) brains. (C) Median fluorescence intensity (MFI) of MeX04 in microglia. (D) MFI of CD45 on microglia. (E) % CD11c+ microglia of total microglia. (F) % MHC-II+ microglia of total microglia. (G) MFI of P2Y12 on microglia. Bars are plotted as mean + SEM. n=5–12. *p<0.05, **p<0.01, ***p<0.001 by two-way ANOVA with Sidak’s multiple comparisons post hoc test.
-
Figure 4—source data 1
Source data associated with Figure 4.
(B) % MeX04+ microglia of total microglia in male and female wild-type (grey), AppNL-G-FCst7+/+ (blue), and AppNL-G-FCst7-/- (red) brains. (C) Median fluorescence intensity (MFI) of MeX04 in microglia. (D) MFI of CD45 on microglia. (E) % CD11c+ microglia of total microglia. (F) % MHC-II+ microglia of total microglia. (G) MFI of P2Y12 on microglia.
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig4-data1-v1.xlsx
The benefit of using a FACS-based approach to investigating microglial Aβ burden is the opportunity to include surface phenotypic markers to assess microglial reactivity. We assessed expression of CD45, a broad but sensitive marker of activation intensity within microglia; P2Y12, a microglial homeostatic marker; CD11c, a marker shown to be upregulated in disease/damage-associated microglia (Keren-Shaul et al., 2017) and MHCII, a marker of antigen presentation, a process which is also dependent on peptide processing through the endolysosomal system (Figure 2—figure supplement 1). AppNL-G-F microglia had increased expression of CD45, CD11c, and MHCII compared to wild-type counterparts (Figure 4D–F). We did not detect a reduction in P2Y12 expression (Figure 4G). Cst7 deletion did not lead to any significant differences in detection of these markers, although we did note a trend towards increased MHCII+ microglia only in female mice, perhaps consistent with elevated endolysosomal activity and our RNASeq data suggesting an increase in antigen presentation pathway genes specifically in this group.
Cst7 negatively regulates phagocytosis in microglia from female AppNL-G-F mice
Our current data suggest that Cst7 plays a sex-dependent regulatory role on microglia leading to increased endolysosomal gene pathways, region-dependent lysosomal activity, and Aβ burden in female AppNL-G-FCst7-/- microglia. However, in vivo it is challenging to ascertain whether the increase in microglial Aβ is due to an increased uptake or to an impaired degradation. To test this, we conducted in vitro assays on Cst7+/+ and Cst7-/- microglia to examine phagolysosomal and related functions. Initially, we examined microglia from young adult Cst7+/+ and Cst7-/- background-matched mice to understand any baseline differences in the absence of a disease exposure. Unlike constitutively highly expressed microglial genes such as Trem2 where knockout leads to dramatic alterations in in vitro phenotype (Takahashi et al., 2005; Hsieh et al., 2009; Kleinberger et al., 2014; Yeh et al., 2016), Cst7 is expressed at very low levels basally (Sala Frigerio et al., 2019; Figure 1A). Therefore, we reasoned that Cst7 deletion would have limited impact on microglial phenotype at baseline where Cst7 is not highly expressed. Comparing Cst7+/+ and Cst7-/- microglia, we observed no significant difference in uptake of Aβ oligomers, human AD synaptoneurosomes, Staphylococcus aureus bioparticles, or myelin debris (Figure 5—figure supplement 1A–D); degradation of Aβ (Figure 5—figure supplement 1E); or lysosomal hydrolysis measured by the ability to cleave the quencher molecule from DQ-BSA (Figure 5—figure supplement 1F); transcriptional response to LPS, IL-4, or apoptotic neurons (Figure 5—figure supplement 1G–I); cytokine secretion in response to LPS or IL-4 (Figure 5—figure supplement 2) or NLRP3 inflammasome activation by silica particles (Figure 5—figure supplement 1J and K). We also addressed the possibility of compensation in adult knockout mice by using siRNA to knock down Cst7 in the BV-2 microglia cell line and found that, despite 70% reduction in gene expression (Figure 5—figure supplement 3A), Cst7 knockdown had no effect on IL-6 secretion (Figure 5—figure supplement 3B) or transcriptional changes response to LPS or IL-4 (Figure 5—figure supplement 3C–E).
Therefore, in order to investigate CF function in vitro in the context of the disease-associated marked induction of microglial Cst7 expression (see above), we developed a model system using primary microglia isolated from 12-month AppNL-G-F mice and cultured them for 3 days in vitro (Figure 5A). We found that microglia isolated from AppNL-G-F mice then cultured for 3 days express 20-fold more Cst7 than from age-matched wild-type counterparts (Figure 5B). We also found that these AppNL-G-F microglia express two-fold less Tmem119, an established microglial homeostatic gene, than wild-type cells (Figure 5C). These data suggest that, despite removal from a disease brain and subsequent culture in vitro, 12-month-old AppNL-G-F microglia retain key elements of their in vivo disease transcriptional identity.
Figure 5 with 4 supplements see all
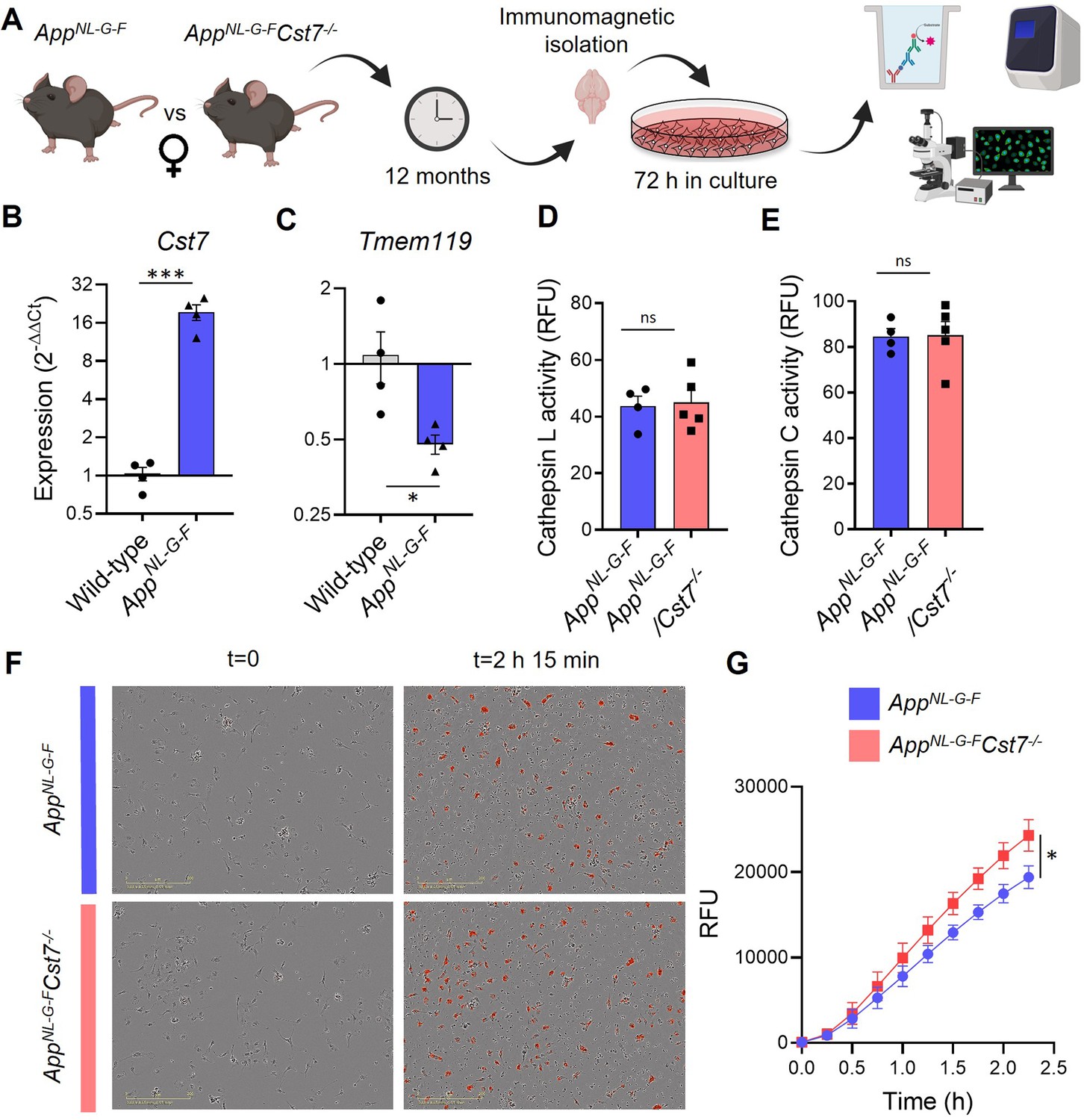
Cst7 negatively regulates phagocytosis in microglia from female AppNL-G-F mice.
(A) Study design schematic. (B–C) qPCR data for Cst7 (B) and Tmem119 (C) RNA expression from female wild-type and AppNL-G-F microglia isolated by CD11b beads and cultured for 3 days in vitro. Bars represent mean fold change calculated by ΔΔCt. n=4. *p<0.05, ***p<0.001 by two-way t-test. (D–E) Cathepsin L (D) and C (E) activity from female AppNL-G-FCst7+/+ (blue) and AppNL-G-FCst7-/- (red) microglia measured by probe-based assay. Bars represent mean relative fluorescence units (RFU) + SEM. n=4–5. (F) Example images from myelin phagocytosis assays with female AppNL-G-FCst7+/+ (blue) and AppNL-G-FCst7-/- (red) microglia taken immediately before (t=0) and 2 hr 45 min after (t=2 hr 45 min) addition of pHrodo-tagged myelin. (G) Quantification of (F). Data shown are mean RFU (calculated as total integrated intensity of pHrodo Red normalised to cell confluence) ± SEM. n=4. *p<0.05 calculated by unpaired t-test on area under curve (AUC) of data.
-
Figure 5—source data 1
Source data associated with Figure 5.
(B–C) qPCR data for Cst7 (B) and Tmem119 (C) RNA expression from female wild-type and AppNL-G-F microglia isolated by CD11b beads and cultured for 3 days in vitro. (D–E) Cathepsin L (D) and C (E) activity from female AppNL-G-FCst7+/+ (blue) and AppNL-G-FCst7-/- (red) microglia measured by probe-based assay. (G) Quantification of myelin phagocytosis assays with female AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- microglia taken immediately before (t=0) and 2 hr 45 min after (t=2 hr 45 min) addition of pHrodo-tagged myelin.
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig5-data1-v1.xlsx
We then used this validated system to compare functional properties of microglia isolated from female AppNL-G-FCst7-/- and AppNL-G-FCst7+/+ mice. First, we showed that Cst7 knockout had no effect on isolation yield (Figure 5—figure supplement 4A) and observed no differences in inflammatory cytokine secretion in response to LPS or NLRP3 activation stimulus silica (Figure 5—figure supplement 4B–D), as predicted from our previous RNASeq analysis in female mice. Next, we tested whether Cst7 plays a role in endolysosomal processes such as phagocytosis or lysosomal degradation. We used a pulse-chase live-imaging assay and found that Cst7 deletion does not affect degradation of Aβ1-42 (Figure 5—figure supplement 4E), a process dependent on cathepsins as it was inhibited by broad-spectrum inhibitors K777 and Ca074-Me (Figure 5—figure supplement 4F). We also showed that Cst7 deletion does not affect intracellular activity of cathepsins L and C (Figure 5D and E), targets of cystatin-F (Hamilton et al., 2008), or microglial lysosomal hydrolysis (Figure 5—figure supplement 4G). These data would suggest that the increased microglial Aβ burden in vivo observed in Figure 4A is not due to decreased degradative capacity and may be due increased phagocytosis. To test this, we performed uptake/phagocytosis assays on microglia isolated from 12-month female AppNL-G-FCst7-/- vs. AppNL-G-FCst7+/+. To remove the confounding factor that female AppNL-G-FCst7-/- will already contain more Aβ (Figure 4B and C) and this may influence further Aβ uptake, we used myelin debris tagged with the pH-sensitive dye pHrodo Red. We found that Cst7 deletion led to a significant increase in microglial phagocytosis of myelin debris (Figure 5F and G). We also tested uptake with fluorescently tagged Aβ1-42 and found a trend towards increased uptake of Aβ1-42 (Figure 5—figure supplement 4H).
Together, these data suggest that, in females, Cst7/CF negatively regulates phagocytosis but does not affect lysosomal proteolysis or PRR-driven inflammatory cytokine production in microglia from the AppNL-G-F model of amyloid-driven AD.
Effect of Cst7 deletion on disease pathology
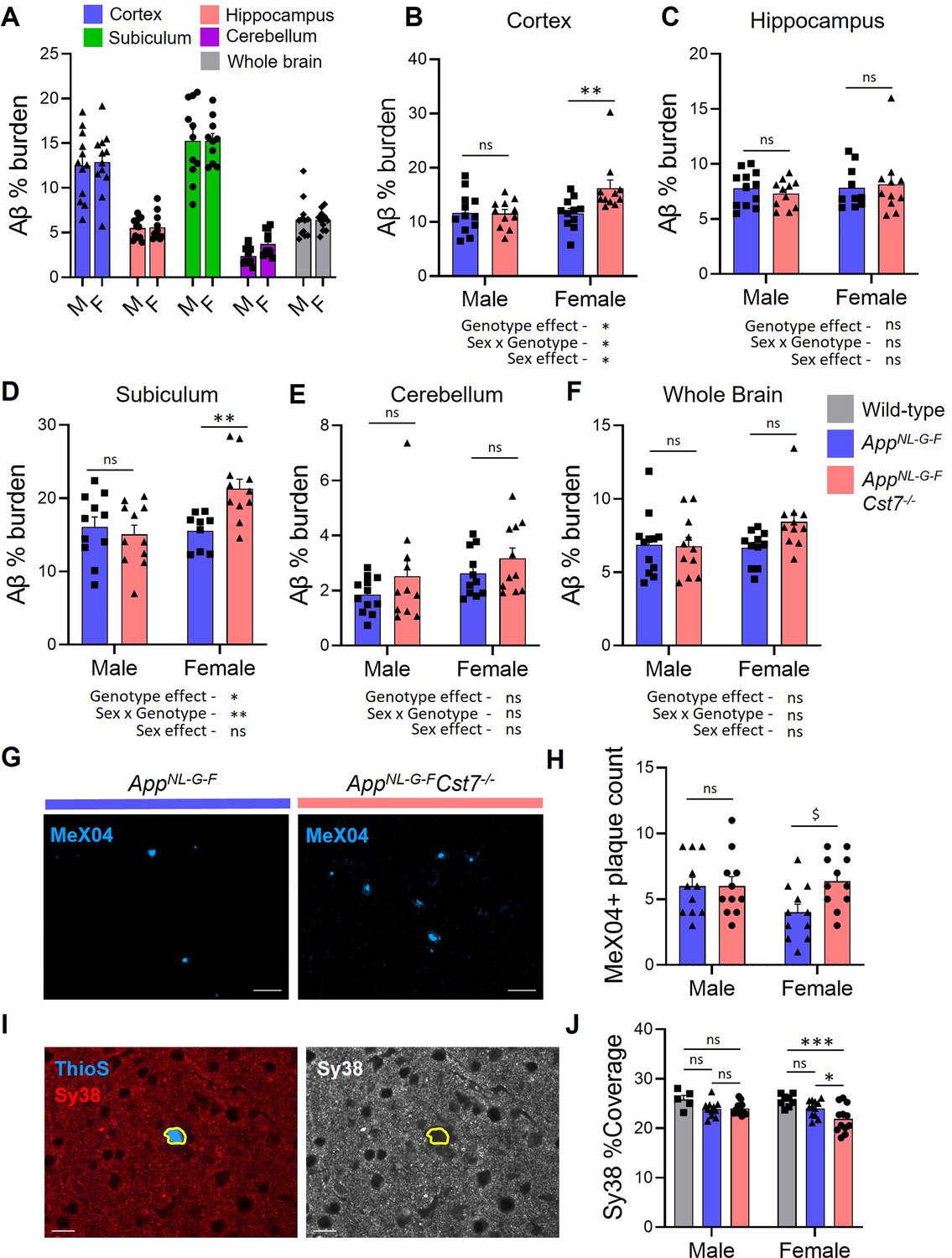
Having discovered that Cst7 plays sex-dependent regulatory roles on microglial phenotype and function in the AppNL-G-F model of amyloid-driven AD, we sought to determine if this is associated with effects on disease pathology. Therefore, we studied the Aβ plaques in 12-month male and female AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- mice across brain regions. We observed the greatest plaque burden in the cortex and dorsal subiculum followed by the hippocampus (Figure 6A) and, as expected, the cerebellum was largely devoid of plaque pathology. Next, we compared plaque burden within each area between AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- male and female mice. In males, there was no effect of Cst7 genotype on plaque burden in any region (Figure 6B–F). Surprisingly, despite an increased lysosomal burden (Figure 3F) and intramicroglial Aβ load (Figure 4B) in female AppNL-G-FCst7-/- mice, there was no decrease in amyloid plaque burden in any region. In fact, the reverse pattern was evident in some areas, most notably in the subiculum (Figure 6D). In order to further probe the increase in amyloid pathology in the subiculum, we used MeX04 labelling of the dense cores of Aβ plaques and counted the number of plaques in this region. We observed a significant although modest increase in plaque number specifically in the females (Figure 6G, quantification in Figure 6H) without any difference in average plaque size detected (Figure 6—figure supplement 1A). Together, these data suggest that Cst7/CF plays a negative regulatory role on microglial endolysosomal function in female AD mice and show that removal of this block and resultant increase in phagocytosis in AppNL-G-FCst7-/- females is associated with increased plaque pathology. Finally, we assessed whether Cst7 deletion had any effect on gross synapse coverage using synaptophysin (Sy38) immunostaining and thioflavin S (ThioS) to localise plaques. We observed loss of Sy38 coverage around plaques (Figure 6I) and a small but significant decrease in coverage between AppNL-G-F/Cst7-/- vs. AppNL-G-F brains only in females (Figure 6J). This reflects the effect observed with plaque coverage above suggesting the increased plaque burden in Cst7-/- female mice may lead to increased synapse loss.
Figure 6 with 1 supplement see all
Investigating the effect of Cst7 deletion on disease pathology.
(A) % Aβ coverage measured by 6E10 3,3'-diaminobenzidine (DAB) staining in cortex (blue), hippocampus (red), subiculum (green), cerebellum (magenta), and whole brain (grey) of male and female AppNL-G-F brains. (B–F) % Aβ coverage in the cortex (B), hippocampus (C), subiculum (D), cerebellum (E), and whole brain (F) of male and female AppNL-G-FCst7+/+ (blue) and AppNL-G-FCst7-/- (red) mice. Points are measured by thresholding of whole region area in sagittal section. Bars represent mean + SEM % coverage. n=10–12. *p<0.05, **p<0.01 calculated by two-way ANOVA with Sidak’s multiple comparisons post hoc test. (G) Example images from subicula of female AppNL-G-FCst7+/+ (left) and AppNL-G-FCst7-/- (right) mice stained with MeX04 (blue). Scale bars are 50 μm. (H) Quantification of (G). MeX04+ plaque count in the subicula of female AppNL-G-FCst7+/+ (blue) and AppNL-G-FCst7-/- (red) mice. Points are number of plaques from a single field of view for each mouse. Bars are mean plaque count + SEM. n=10–12. $p<0.05 calculated by two-way Mann-Whitney test. (I) Example image of a plaque with Thio S (blue) and synapses stained with synaptophysin (red) in AppNL-G-F brain. Plaque border is highlighted in yellow. (J) Quantification of (I) measuring coverage of synaptophysin. Bars represent mean + SEM % coverage. n=5–12. *p<0.05 calculated by two-way ANOVA with Tukey’s multiple comparisons post hoc test.
-
Figure 6—source data 1
Source data associated with Figure 6.
(A) % Aβ coverage measured by 6E10 3,3'-diaminobenzidine (DAB) staining in cortex, hippocampus, subiculum, cerebellum, and whole brain of male and female AppNL-G-F brains. (B–F) % Aβ coverage in the cortex (B), hippocampus (C), subiculum (D), cerebellum (E), and whole brain (F) of male and female AppNL-G-FCst7+/+ and AppNL-G-FCst7-/- mice. Points are measured by thresholding of whole region area in sagittal section. (H) Quantification of MeX04+ plaque count in the subicula of female AppNL-G-FCst7+/+ and AppNL-G-FCst7-/-. (J) Quantification of % coverage of synaptophysin in the cortex of male and female wild-type, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- brains.
- https://cdn.elifesciences.org/articles/85279/elife-85279-fig6-data1-v1.xlsx
Discussion
Changes in microglial cells have been heavily implicated in the pathogenesis of AD, particularly supported by genetics studies, where expression of over 50% of risk genes is enriched in microglia (Hansen et al., 2018). Notably, a transcriptomic profile has emerged involving Trem2-dependent downregulation of homeostatic genes and upregulation of a signature characterised by genes involved in phagocytosis, lipid handling, and endolysosomal transport (Deczkowska et al., 2018). One of the most robustly upregulated of these genes, Cst7 (CF), is believed to be involved in protease inhibition. However, the cellular functions controlled by Cst7/CF in microglia and whether Cst7/CF has a disease-modifying role in AD-related CNS proteinopathy has not been explored previously. Here, we reveal that Cst7/CF plays a sexually dimorphic role in microglia in an amyloid-driven AD model, acting as a restraint on microglial endolysosomal activity and phagocytosis specifically in female mice, that when absent results in subtle aggravation of Aβ pathology and synapse loss.
Since their initial description in 2017, the DAM/MGnD/ARM gene signature has been extensively investigated with over 4000 citations between the three papers describing them in 5 years. However, beyond GO analysis, there is surprisingly little understanding of what many of the hallmark genes comprising this signature functionally do in the context of neurodegenerative disease. This is the first study to our knowledge to test the functional role of Cst7, one of the most consistently replicated markers of this cell state transcriptomic signature, in β-amyloid-driven pathology. Perhaps counterintuitively, Cst7 (a reported cysteine protease inhibitor) is robustly upregulated in microglia alongside concomitant upregulation of cysteine proteases such as cathepsins D, B, L, and Z. In fact, such internal regulation systems are not uncommon in inflammation biology, where proinflammatory mediators such as IL-1β and IL-18 are negatively regulated by co-expressed IL-1 receptor antagonist and IL-18 binding protein, respectively (Hurme and Santtila, 1998; Dinarello et al., 2013). We therefore hypothesise that Cst7/CF is upregulated along with numerous cathepsin genes in order to prevent potentially harmful protease activity (Turk et al., 2012). In this study, we show that Cst7/CF knockout triggers an increase in microglial lysosomal activity and amyloid uptake. Intriguingly, this did not seem to depend on overactive intracellular cathepsin activity, which intuitively would have been expected to increase with the lack of negative regulator. It is possible that CF deficiency does in fact lead to an increase in cathepsin activity, but CF itself is secreted and only impairs secreted cathepsins which are responsible for promoting phagocytosis. Indeed, studies have suggested that CF can be secreted and that there is a role for secreted cathepsins in phagocytosis (Liuzzo et al., 1999; Hamilton et al., 2008). This is also consistent with the observation that exogenously applied cystatin C can block phagocytosis in human polymorphonuclear neutrophils (Leung-Tack et al., 1990).
Another key implication from this study is that of sexual dimorphism in disease. Here, we show stark differences in the effect of Cst7 deletion between males and females at both the gene and protein level. Although there is a relative paucity of studies investigating male vs. female microglia in homeostasis and disease, numerous differences have been identified (Lynch, 2022). In general, male microglia are believed to have higher baseline activity in processes such as inflammation, antigen presentation, and phagocytosis; whereas female microglia are more associated with neuroprotection and the DAM/ARM/MGnD signature in ageing and disease (Guneykaya et al., 2018; Kodama and Gan, 2019; Sala Frigerio et al., 2019; Villa et al., 2019; Guillot-Sestier et al., 2021). These differences in baseline properties are only partially dependent on hormonal signalling as masculinisation of females by ovariectomy and hormone replacement only partially recapitulated male microglia (Villa et al., 2018). Indeed, sexual dimorphism of microglia in adults appears a cell-intrinsic phenomenon as female microglia transplanted in male brains retain their transcriptional profile and reduce infarct size in males after ischemic stroke (Villa et al., 2018). Interestingly, we generally do not observe marked sexual dimorphism in microglia in wild-type or AppNL-G-F mice. However, DAM/ARM/MGnD gene expression was higher in female vs. male microglia and lysosomal activity was markedly higher in male vs. female brains measured by LAMP2 staining, which is consistent with the literature (Guneykaya et al., 2018; Sala Frigerio et al., 2019). Importantly, we observed a substantial interaction effect of Cst7 deletion and sex, whereby removal of the Cst7 gene led to an ‘unlocking’ of sexual dimorphism in our cohort. This is most clearly demonstrated by DEGs between male vs. female microglia rising from 33 in AppNL-G-FCst7+/+ mice to 240 in AppNL-G-FCst7-/- mice. This sex-dependent interaction effect has previously been observed in studies investigating the cystatin/cathepsin system, where deletion of Cst3 (cystatin C) led to protection in the EAE model of multiple sclerosis in females but not males (Hoghooghi et al., 2020). This effect appeared to be sensitive to hormones as ovariectomy or castration followed by testosterone or estrogen/progesterone administration effectively reversed the effect of Cst3 knockout. The precise molecular mechanism underpinning this was not explored but could include differential post-translational modification. The inhibitor effect of cystatin C on phagocytosis is dependent on N-terminal truncation (Leung-Tack et al., 1990) whilst more recently an N-terminal processing event has been identified for CF (Hamilton et al., 2008). Understanding whether N-terminal processing events are sex-specific would be a useful step in determining how genetic deletion can have differential effects dependent on sex. Indeed, sex:genetics interactions are not uncommon in neurodegenerative disease, with various risk variants posing greater or lesser effect on risk in men or women (Gamache et al., 2020). One example of this interaction is APOE, in which the presence of the E4 allele confers a greater AD risk in women than in men (Altmann et al., 2014).
Ideas and speculation
Here, we observed that Cst7/CF played a role in microglial amyloid uptake and endolysosomal gene expression only in females. Surprisingly, this did not lead to a decrease in Aβ plaque pathology as might be expected but an increase, although of modest degree, in specific brain areas. This discovery raises the important question of the relationship between microglial phagocytosis and plaques. It is generally considered that an increase in microglial phagocytosis should lead to reduction in plaque burden and benefit in disease (Heneka et al., 2013; Tejera et al., 2019). However, recent studies have emerged to suggest that phagocytosing microglia may in fact act to ‘build’ plaques rather than dismantle them. For example, microglia lacking TAM receptors Axl and Mer fail to take up Aβ which leads to an overall decrease in dense-core plaque formation (Huang et al., 2021), microglia can ‘seed’ plaques from disease tissue into engrafted, non-affected regions (d’Errico et al., 2022), and either genetic or pharmacological depletion of microglia leads to a reduction in plaque burden/intensity accompanied by increase in cerebral amyloid angiopathy (Spangenberg et al., 2019; Kiani Shabestari et al., 2022) perhaps through redistributing Aβ. Interestingly, only female mice were used in two of these studies (Huang et al., 2021; d’Errico et al., 2022), which parallels our sex-specific findings with Cst7. In the above studies, microglial depletion/inactivation leading to reduced parenchymal plaque burden is generally detrimental in disease. In contrast, we find indications in our study that the increased microglial phagocytosis and concomitant increase in plaque burden in Cst7-/- females may be detrimental. This is most likely due to direct effect of increased plaque burden leading to increased synapse loss. However, we cannot confirm whether this apparent synapse loss is a result of increased microglial uptake (Hong et al., 2016) or direct synaptotoxicity (Tzioras et al., 2023b). We speculate that Cst7/CF may contribute to restraining plaque formation and protecting against synapse loss by influencing microglial endolysosomal function in a dose-dependent manner.
We were also surprised that there was no change in plaque burden in males, where microglia had a reduction in inflammatory profile. Studies have suggested that reducing inflammatory cytokine secretion may reduce plaque formation and increase Aβ uptake (Heneka et al., 2013; Tejera et al., 2019). These effects may be linked specifically to the NLRP3-caspase-1-ASC-IL-1β axis and that a more general reduction in inflammatory genes is not capable of phenocopying Nlrp3-/- mice. Additionally, we observe a relatively mild reduction in inflammatory genes in male AppNL-G-FCst7-/- mice with fold changes mostly below 2 vs. AppNL-G-FCst7+/+ controls. We speculate that either complete genetic deletion or strong inhibition with chemical compounds is required to yield pro-phagocytic effects. We also note in our study that male Cst7-/- microglia had reduced expression of lysosomal proteins, possibly counteracting the reduction in inflammatory gene expression.
Limitations
Although these data provide novel insight into the sex-dependent role of microglial gene Cst7 in AD, it is important to acknowledge some caveats of the study that point to potential future investigations into this complex biological phenomenon. Here, we investigated CF function solely in mouse. While human data on microglial CF is limited, studies have shown that (where detected) CF is expressed predominantly in myeloid cells (Zhang et al., 2016; Mathys et al., 2019; Schirmer et al., 2019; Srinivasan et al., 2020) and identified CF expression in microglia around plaques in AD (Ofengeim et al., 2017) and CST7 enrichment in AD-associated microglia clusters by single nucleus RNA sequencing (snRNASeq) (Gerrits et al., 2021). CST7 protein has also been detected as higher levels predicted slower tau accumulation and cognitive decline with a significant sex interaction effect (Pereira et al., 2022). However, CF/CST7 has also remained undetected in similar immunostaining and snRNASeq studies (Nuvolone et al., 2017; Zhou et al., 2020). While the reason for this discrepancy is unclear, it may be partially due to sex. Indeed, there was a greater ratio samples from female brains in Gerrits et al., 2021 than in Zhou et al., 2020, as would be expected if CST7 plays a more important role in female microglia. These data further demonstrate the importance of stratifying studies by sex. Additionally, while mechanistic studies investigating CF/CST7 in human microglia differentiated from inducible pluripotent stem cells would be valuable, our discovery that CF function is revealed in mouse microglia only when cells are taken from disease context would suggest more complex models are required. Indeed, a recent study utilised overexpression of TREM2 in human microglial cells to show that driving CST7 expression led to inhibition of phagocytosis in a CST7-dependent manner (Popescu et al., 2023). These data suggest the functional role of CF is not species-specific and provide important translational evidence. Finally, while this study details the discovery of the mechanistic role of CF/Cst7 in an amyloid-driven AD model, the precise mechanism(s) by which Cst7 deletion affects microglia only in females and the biological reason for the accelerated DAM/MGnD/ARM programme in females now demonstrated in multiple studies remains unknown. This phenomenon may go some way to explaining the sexual dimorphism we observe in this study, that is, the effect of Cst7 deletion at 12 months in females may be mirrored in males at later disease stages. However, although we do observe increased expression of sex-regulated DAM genes such as Gpnmb and Spp1, we do not find differential expression of Cst7 itself in females vs. males or most other genes in AppNL-G-F. Additionally, we find a qualitatively different pattern of altered gene expression from Cst7 deletion in female vs. male AppNL-G-F mice rather than similar effects of differing magnitude. This suggests that that a sex-intrinsic mechanism rather than a simple ‘lagging’ in males is a more likely explanation underpinning Cst7 deletion sex-dependent effects. Future work should investigate this further with ovariectomy/hormone replacement and transplantation studies, such as those performed by Villa et al., 2018.
In summary, our data provide key mechanistic insight into the role of one of the most robustly upregulated genes in disease-reactive microglia. These data suggest that Cst7/CF regulates some key aspects of microglial function, in a sex-dependent manner, and that these are associated with pathology-influencing effects in an amyloid-driven AD model that also manifest differently in males and females. We hypothesise that Cst7/CF plays a part in an internal regulatory system that balances the two crucial processes of phagocytosis and inflammatory signalling, which are sexually dimorphic processes. More broadly, and in view of the poorly understood functions of many other recently described hallmark microglial disease-related state mediators, it is imperative to consider interactions with sex when investigating their cellular and disease roles.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (Mus musculus) | AppNL-G-F | DOI:10.1038/nn.3697 | ||
| Genetic reagent (Mus musculus) | Cst7-/- | DOI:10.1016/j.immuni.2016.03.003 | ||
| Cell line (Mus musculus) | BV2 | Collaborator | RRID: CVCL_0182 | Authenticated (STR profiling) |
| Cell line (Homo sapiens) | SH-SY5Y | Collaborator | RRID: CVCL_0019 | Authenticated (STR profiling) |
| Biological sample (Homo sapiens) | Synaptoneurosome from AD brain | DOI:10.1016/j.xcrm.2023.101175 | AMREC (approval number 15-HV-016) | |
| Biological sample (Mus musculus) | Purified myelin | This paper | See Materials and methods ‘Myelin purification and pHrodo tagging’ | |
| Antibody | Anti-Ly6C- Alexa Fluor 488 (Rat monoclonal) | BioLegend | Cat# 128021 | FACS 1:500 |
| Antibody | Anti-P2Y12 – PE (Rat monoclonal) | BioLegend | Cat# 848003 | FACS 1:200 |
| Antibody | Anti-MHCII – PEDazzle594 (Rat monoclonal) | BioLegend | Cat# 107647 | FACS 1:200 |
| Antibody | Anti-CD45 – PE-Cy7 (Rat monoclonal) | BioLegend | Cat# 103113 | FACS 1:200 |
| Antibody | Anti- CD11c – APC (Armenian Hamster monoclonal) | BioLegend | Cat# 117309 | FACS 1:200 |
| Antibody | Anti- CD11b – BV711 (Rat monoclonal) | BioLegend | Cat# 848003 | FACS 1:50 |
| Antibody | Anti-IBA1 (Rabbit polyclonal) | Wako | Cat# 019-19741 | IF 1:2000 |
| Antibody | Anti- Aβ (6E10) (Mouse monoclonal) | BioLegend | Cat# 803001 | IF/DAB 1:1000 |
| Antibody | Anti-LAMP2 (Rat monoclonal) | BioLegend | Cat# 108501 | IF 1:200 |
| Antibody | Anti-Synaptophysin (Sy38) (Mouse monoclonal) | Abcam | ab8049 | IF 1:200 |
| Antibody | Anti-rabbit IgG Alexa Fluor 488 (Goat polyclonal) | ThermoFisher | Cat# A-11008 | IF 1:500 |
| Antibody | Anti-mouse IgG Alexa Fluor 555 (Donkey polyclonal) | ThermoFisher | Cat# A-31570 | IF 1:500 |
| Antibody | Anti-rat IgG Alexa Fluor 647 (Goat polyclonal) | ThermoFisher | Cat# A-21247 | IF 1:500 |
| Antibody | Anti-mouse Biotinylated | Vector Labs | Cat# ba-9200 | DAB 1:100 |
| Chemical compound, drug | Methoxy X04 | Tocris | Cat# 4920 | |
| Chemical compound, drug | Cathepsin L probe | Bachem | Cat# 4003379 | Z-Phe-Arg-AMC |
| Chemical compound, drug | Cathepsin C probe | Bachem | Cat# 4003759 | H-Gly-Phe-AMC |
| Sequence-based reagent | Cst7_F | This paper | PCR primers | ACCAATAACCCAGGAGTGCTTA |
| Sequence-based reagent | Cst7_R | This paper | PCR primers | TGACCCAGACTTCAGAGTAGCA |
| Sequence-based reagent | Arg1_F | This paper | PCR primers | GGAGACCACAGTCTGGCAGTTGGA |
| Sequence-based reagent | Arg1_R | This paper | PCR primers | GGACACAGGTTGCCCATGCAGA |
| Sequence-based reagent | Il1b_F | This paper | PCR primers | CGACAAAATACCTGTGGCCTTGGGC |
| Sequence-based reagent | Il1b_R | This paper | PCR primers | TGCTTGGGATCCACACTCTCCAGC |
| Sequence-based reagent | Trem2_F | This paper | PCR primers | CTGCTGATCACAGCCCTGTCCCAA |
| Sequence-based reagent | Trem2_R | This paper | PCR primers | CCCCCAGTGCTTCAAGGCGTCATA |
| Sequence-based reagent | Gapdh_F | This paper | PCR primers | TGCATCCACTGGTGCTGCCAA |
| Sequence-based reagent | Gapdh_R | This paper | PCR primers | ACTTGGCAGGTTTCTCCAGGCG |
| Sequence-based reagent | Lilrb4a_F | This paper | PCR primers | ATGGGCACAAAAAGAAGGCTAA |
| Sequence-based reagent | Lilrb4a_R | This paper | PCR primers | GGCATAGGTTACATCCTGGGTC |
| Sequence-based reagent | Ndufv1_F | This paper | PCR primers | ATTTTCTCGGCGGGTTGGTT |
| Sequence-based reagent | Ndufv1_R | This paper | PCR primers | CACCTTTCAGCCTCCAGTCA |
| Commercial assay or kit | DuoSet ELISA – IL-1β | R&D Systems | Cat# DY401 | |
| Commercial assay or kit | DuoSet ELISA – IL-6 | R&D Systems | Cat# DY406 | |
| Commercial assay or kit | DuoSet ELISA – TNFα | R&D Systems | Cat# DY411 | |
| Commercial assay or kit | RNAscope 2.5 HD Duplex Assay | Biotechne | Cat# 322436 | |
| Commercial assay or kit | RNAscope probes – Cst7 | Biotechne | Cat# 498711-C2 | |
| Commercial assay or kit | RNAscope probes – Lilrb4a | Biotechne | Cat# 1260291-C2 |
Animals
AppNL-G-F mice (Saito et al., 2014) were provided by the RIKEN BRC through the National BioResource Project of the MEXT/AMED, Japan. Cst7-/- mice (Matthews et al., 2016) were obtained from Colin Watts, University of Dundee, UK. C57Bl/6J mice used for myelin preparations were purchased from Charles River, UK. Animals were maintained under standard laboratory conditions: ambient temperatures of 21°C (±2°C), humidity of 40–50%, 12 hr light/dark cycle, ad libitum access to water, and standard rodent chow. Genotype groups were randomised during the study and experimenters were blinded to genotype during all experiments. All animal experiments were carried out in accordance with the United Kingdom Animals (Scientific Procedures) Act 1986 and approved by the Home Office and the local Animal Ethical Review Group, University of Edinburgh. Experimental design, analysis, and reporting followed the ARRIVE 2.0 guidelines (Percie du Sert et al., 2020). Genotyping was performed using optimised assays from Transnetyx. Methoxy X-04 (MeX04, BioTechne) was reconstituted in DMSO at 10 mg/mL before diluting to 0.33 mg/mL in 6.67% Cremaphor EL (Fluka), 90% PBS (Merck). MeX04 was administered by intraperitoneal injection at 10 mg/kg 2.5 hr before animals were terminated.
Microglial FACS isolation
Request a detailed protocolMicroglia were isolated by enzymatic digestion and FACS as follows. Brains from 12-month-old male and female AppWt/WtCst7+/+, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice were isolated by terminally anaesthetising with 3% isoflurane (33.3% O2 and 66.6% N2O) and transcardial perfusion with ice-cold DEPC-treated 0.9% NaCl, 0.4% trisodium citrate. Brains were immediately separated down the midline then the right hemisphere placed into ice-cold HBSS (ThermoFisher) and minced using a 22A scalpel. Minced hemi-brains were then centrifuged (300 × g, 2 min) and digested using the MACS Neural Dissociation Kit (Miltenyi) according to the manufacturer’s instructions. Briefly, brain tissue was incubated in enzyme P (50 μL/hemi-brain) diluted in buffer X (1900 μL/hemi-brain) for 15 min at 37°C under gentle rotation before addition of enzyme A (10 μL/hemi-brain) in buffer Y (20 μL/hemi-brain) and further incubation for 20 min at 37°C under gentle rotation. Following digestion, tissue was dissociated mechanically using a Dounce homogeniser (loose pestle, 20 passes) on ice and centrifuged (400 × g, 5 min at 4°C). To remove myelin, tissue was resuspended in 35% isotonic Percoll (GE Healthcare) in HBSS overlaid with 1× HBSS and centrifuged (800 × g, 30 min, 4°C, no brake). Following centrifugation, the supernatant and myelin layers were discarded and the pellet resuspended in 100 μL FACS buffer (PBS, 0.1% low endotoxin bovine serum albumin [BSA, Merck], 25 mM HEPES) and transferred to V-bottomed 96-well plates (ThermoFisher). Cell suspensions were treated with anti-mouse-CD16/32 to block Fc receptors (BioLegend, 5 μg/mL, 20 min at 4°C) before washing and transferring to PBS and treating with Zombie NIR (BioLegend, 1:100, 15 min at room temperature [RT]) to label dead cells. Next, cells were stained with antibodies (Table 1 ) to identify microglia and incubated for 20 min at 4°C. Antibodies (BioLegend) used were as follows.
Table 1
Antibodies used for flow cytometry.
| Fluorochrome | Antigen | Clone | Lot | Cat number | Stock concentration (mg/mL) | Dilution |
|---|---|---|---|---|---|---|
| Alexa Fluor 488 | Ly6C | HK1.4 | B248739 | 128021 | 0.5 | 500 |
| PE | P2Y12 | S16007D | B298459 | 848003 | 0.2 | 200 |
| PEDazzle594 | MHCII | M5/114.15.2 | B216062 | 107647 | 0.2 | 200 |
| PE-Cy7 | CD45 | 30-F11 | B271123 | 103113 | 0.2 | 200 |
| APC | CD11c | N418 | B280313 | 117309 | 0.2 | 200 |
| BV711 | CD11b | M1/70 | B305911 | 101241 | 0.005 | 50 |
Following incubation, cells were washed with FACS buffer, resuspended in 500 μL FACS buffer and transferred to 5 mL round-bottom FACS tubes through cell strainer caps (BD). Cells were sorted through a 100 μm nozzle on a FACS Aria II (BD) at the QMRI Flow Cytometry Cell Sorting Facility, University of Edinburgh with the following strategy (Figure 2—figure supplement 1). Debris were eliminated using forward scatter (area) vs. side scatter (area), singlets were isolated using forward scatter (area) vs. forward scatter (height), dead cells were eliminated with Zombie NIR, monocytes were eliminated by gating out Ly6C+ cells, and microglia were defined as CD11b+/CD45+. All antibodies were validated using appropriate fluorescence minus one controls. Microglia were sorted directly into 500 μL RLT buffer (QIAGEN) and 20,000 CD11b+/CD45+ events were collected for downstream analysis using FlowJo software.
RNA purification, QC, and sequencing
Request a detailed protocolRNA was purified using RNEasy Plus Micro kits (QIAGEN) according to the manufacturer’s instructions. RNA was quantified and QC’d using a 4200 TapeStation System (Agilent) with a High Sensitivity RNA ScreenTape Assay according to the manufacturer’s instructions. For in vivo experiments, RNA was pooled from three to four animals within the same experimental group. RNA within pools was equally distributed in amount between the animals. One ng of total RNA was taken forward for low-input RNA sequencing (Cambridge Genomics). Briefly, cDNA was generated using TakaraBio SMART-Seq v4 Ultra Low Input RNA kit before input into the Illumina Nextera XT library prep. Pooled libraries were sequenced on the NextSeq 500 (Illumina) using the 75 cycle High Output sequencing run kit, spiked with 5% PhiX at a depth of 30 million reads per sample.
RNASeq analysis
Request a detailed protocolReads were mapped to the mouse primary genome assembly GRCm39, Ensembl release 104 (Cunningham et al., 2022) using STAR version 2.7.9a (Dobin et al., 2013), and tables of per-gene read counts were summarised using featureCounts version 2.0.2 (Liao et al., 2014). Differential expression analysis was performed using DESeq2 (R package version 1.30.1) (Love et al., 2014), using an adjusted p-value cut-off of 0.05 to identify genes differentially expressed between conditions. Comparisons carried out were male (M) AppWt/WtCst7+/+ vs. M AppNL-G-FCst7+/+, female (F) AppWt/WtCst7+/+ vs. F AppNL-G-FCst7+/+, M AppNL-G-FCst7+/+ vs. M AppNL-G-FCst7-/-, F AppNL-G-FCst7+/+ vs. F AppNL-G-FCst7-/-, M AppNL-G-FCst7+/+ vs. F AppNL-G-FCst7+/+, M AppNL-G-FCst7-/- vs. F AppNL-G-FCst7-/-. For each comparison GO enrichment analysis was performed to provide insight into the biological pathways and processes affected; GO enrichment analysis was performed using topGO (Alexa et al., 2006) (R package version 2.42.0). Significantly upregulated and downregulated gene sets were compared against annotated GO terms from the categories of ‘biological process’. Selected statistically significant GO:BP terms were then visualised using GraphPad prism v9.
qPCR
Request a detailed protocolCells were seeded at 75,000 cells/well in a 24-well plate 7 days before stimulation (primary microglia) or 250,000 cells/well in a 24-well plate 18 hr before stimulation (BV-2 cells). On the day of the assay, microglia were treated with vehicle (PBS, 24 hr, Merck), LPS (100 ng/mL, 24 hr, Merck), IL-4 (20 ng/mL, 24 hr, R&D Systems) or apoptotic SH-SY5Y neuroblastoma cells (2:1 ratio neurons:microglia, 24 hr) then lysed in RLT buffer and RNA isolated according to the manufacturer’s instructions. RNA (100–500 ng) was converted to cDNA using SuperScript IV Reverse Transcriptase (ThermoFisher) according to the manufacturer’s instructions. qPCR was performed using PowerUp SYBR Green PCR Master Mix (ThermoFisher) in 384-well format using an qTOWER384 Real-time PCR machine (Analytik Jena). For BV-2 cells, 10 ng cDNA (assuming 100% RNA to cDNA conversion) was loaded per well with 5 pmol primer/well in triplicate in a total volume of 10 μL. For primary microglia experiments, 0.5 ng cDNA was loaded per well. Data were normalised to the expression of the housekeeping gene Gapdh or mean Gapdh and Ndufv1 and were analysed using the ΔΔCt method. Primers used are detailed in Table 2.
Table 2
Primers used for qPCR.
| Gene | Forward | Reverse |
|---|---|---|
| Cst7 | ACCAATAACCCAGGAGTGCTTA | TGACCCAGACTTCAGAGTAGCA |
| Arg1 | GGAGACCACAGTCTGGCAGTTGGA | GGACACAGGTTGCCCATGCAGA |
| Il1b | CGACAAAATACCTGTGGCCTTGGGC | TGCTTGGGATCCACACTCTCCAGC |
| Trem2 | CTGCTGATCACAGCCCTGTCCCAA | CCCCCAGTGCTTCAAGGCGTCATA |
| Gapdh | TGCATCCACTGGTGCTGCCAA | ACTTGGCAGGTTTCTCCAGGCG |
| Lilrb4a | ATGGGCACAAAAAGAAGGCTAA | GGCATAGGTTACATCCTGGGTC |
| Ndufv1 | ATTTTCTCGGCGGGTTGGTT | CACCTTTCAGCCTCCAGTCA |
Microglial bead-based isolation for in vitro studies
Request a detailed protocolPrimary adult mouse microglia were isolated and cultured as described previously (Grabert and McColl, 2018). Brains were isolated by terminally anaesthetising with 3% isoflurane (33.3% O2 and 66.6% N2O) and transcardial perfusion with ice-cold 0.9% NaCl. Brains were immediately placed into ice-cold HBSS (ThermoFisher) and minced using a 22A scalpel before centrifugation (300 × g, 2 min) and digestion using the MACS Neural Dissociation Kit (Miltenyi) according to the manufacturer’s instructions. Briefly, brain tissue was incubated in enzyme P (50 μL/brain) diluted in buffer X (1900 μL/brain) for 15 min at 37°C under gentle rotation before addition of enzyme A (10 μL/brain) in buffer Y (20 μL/brain) and further incubation for 20 min at 37°C under gentle rotation. Following digestion, tissue was dissociated mechanically using a Dounce homogeniser (loose pestle, 20 passes) on ice and centrifuged (400 × g, 5 min at 4°C). To remove myelin, tissue was resuspended in 35% isotonic Percoll (GE Healthcare) overlaid with HBSS and centrifuged (800 × g, 40 min, 4°C). Following centrifugation, the supernatant and myelin layers were discarded and the pellet resuspended in MACS buffer (PBS, 0.5% low endotoxin BSA [Merck], 2 mM EDTA, 90 μL/brain). Anti-CD11b microbeads (Miltneyi) were added (10 μL/brain) and the suspension incubated for 15 min at 4°C before running through pre-rinsed (MACS buffer) LS columns attached to a magnet (Miltenyi). After washing with 12 mL MACS buffer, columns were removed from the magnet and cells retained (microglia) were flushed in 5 mL MACS buffer. For experiments investigating expression of RNASeq hit genes in non-diseased microglia, cells were immediately lysed in RLT buffer and RNA purified as above. For culture experiments, microglia were resuspended in Dulbecco’s Modified Eagle’s Medium/Nutrient Mixture F-12 (DMEM/F-12, ThermoFisher) supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin (PenStrep, Merck), 10% heat-inactivated fetal bovine serum (FBS, ThermoFisher), 50 ng/mL rhTGFβ-1 (Miltenyi), 10 ng/μL mCSF1 (R&D Systems). Microglia were counted using a haemocytometer and plated out onto 24- or 96-well plates (Corning) coated with poly-L-lysine (Merck). Cells were cultured for 7 days with a half media change on day 3. For stimulation experiments, cells were stimulated with LPS, IL-4, or silica particles (US Silica) at concentrations and timepoints as described in each experiment in detail before supernatant was taken and assessed for IL-6, IL-1β, or TNF-α by DuoSet ELISA (R&D Systems) as per the manufacturer’s instructions.
Immortalised cell culture
Request a detailed protocolMurine BV-2 microglial cells (Blasi et al., 1990) were cultured in DMEM (ThermoFisher), 10% FBS (ThermoFisher), and 1% PenStrep (Merck). Human SH-SY5Y neuroblastoma cells (ECACC 94030304) were cultured in DMEM/F-12 (ThermoFisher), 10% FBS (ThermoFisher), and 1% PenStrep (Merck). To induce apoptosis, SH-SY5Y cells were plated in a 10 cm dish at 4×106 cells/dish and stimulated with UV light (4×15 W UV bulb, 30 hr) before incubation at 37°C for 18 hr. After incubation, cells were removed from dishes using Accutase (Merck) and adjusted to 4×106 cells/mL for stimulation onto microglia. All cells were cultured at 37°C, 5% CO2 in a tissue culture incubator (Panasonic) and handled in a Safe 2020 Class II tissue culture cabinet (ThermoFisher). Cell lines (BV-2 immortalized mouse microglia and SH-SY5Y human neuroblastoma) were obtained from collaborators. Identities of the cell lines were authenticated by STR profiling (American Type Culture Collection) and tested negative for mycoplasma (Mycoplasma Detection Kit, Lonza).
BV-2 siRNA knockdown
Request a detailed protocolSmall interfering RNAs (siRNAs) (Silencer-Select, Ambion) targeting Cst7 were used to induce gene knockdown in BV-2 cells. Cells were plated out at 250,000 cells/well in a 24-well plate 18 hr before knockdown. For transfection of each siRNA (Cst7 and non-targeting control), a 1:1 ratio of Lipofectamine RNAiMAX (ThermoFisher) and the siRNA, both diluted in Opti-MEM (ThermoFisher), were added to desired cell supernatants, with 10 pmol siRNA and 1.5 μL Lipofectamine RNAiMAX used per well. After 24 hr, cells were stimulated with vehicle (PBS), LPS (100 ng/mL, 24 hr, Merck), or IL-4 (20 ng/mL, 24 hr, R&D Systems). Following stimulation, cells were lysed for RNA analysis and supernatants were analysed for IL-6 content by ELISA (DuoSet, R&D Systems) according to the manufacturer’s instructions.
Myelin purification and pHrodo tagging
Request a detailed protocolPurified myelin was prepared from C57Bl/6J mouse brains. Brains were isolated, digested, and myelin layer fractionated using 35% percoll gradient as described above. Myelin was washed by dilution in HBSS and centrifuged at 400 × g for 5 min at 4°C before suspending in 3 mL ice-cold 0.32 M sucrose solution containing protease inhibitor cocktail (Roche). Next, myelin was layered onto 3 mL ice-cold 0.85 M sucrose solution in a 10 mL ultracentrifuge tube (Beckman) before centrifugation at 75,000 × g for 30 min at 4°C on a Beckman Ultracentrifuge with MLA-55 rotor (acceleration 7, deceleration 7). Myelin was collected from the 0.32 M:0.85 M interface and placed in 5 mL ice-cold ddH2O in a new Beckman ultracentrifuge tube and vortexed before further centrifugation at 75,000 × g for 15 min at 4°C (acceleration 9, deceleration 9). Myelin was then reconstituted in 5 mL ddH2O and incubated on ice for 10 min before further centrifugation at 12,000 × g for 15 min at 4°C (acceleration 9, deceleration 9). This wash and ddH2O incubation step was repeated before myelin was again fractionated in 0.32 M:0.85 M sucrose gradient and washed as before. Finally, purified myelin was resuspended in 1 mL sterile PBS and stored at –80°C until further use. Myelin was tagged with pHrodo Red according to the manufacturer’s instructions. Briefly, myelin was centrifuged (10,000 × g 10 min) and resuspended in 100 μL pHrodo Red succinimidyl ester (100 μg/mL in PBS 1% DMSO). Myelin was incubated for 45 min at RT in the dark before washing × 2 with 1 mL PBS. Finally, tagged myelin was resuspended in 100 μL, aliquoted, and stored at –20°C for further use.
Microglial phagocytosis/degradation assays
Request a detailed protocolPrimary adult mouse microglia were isolated as described above and plated out in 96-well plates at 50,000 cells/well. Cells were cultured for 7 days or 3 days (in the case of cells from AppNL-G-F brains) before stimulation. For phagocytosis assays, microglia were first imaged using an IncuCyte S3 Live-Cell Analysis System (Sartorius) in ‘phase’ and ‘red’ channels to gain baseline information on confluence and background fluorescence. Next, cells were stimulated with Aβ1-42HiLyte647 (ThermoFisher, 0.5 μM), pHrodo Red S. aureus Bioparticles (ThermoFisher, 100 μg/mL), purified mouse myelin tagged with pHrodo Red succinimidyl ester (1:100, ThermoFisher), or isolated human synaptoneurosomes from AD brains (Tai et al., 2014; Tzioras et al., 2023a) tagged with pHrodo Red succinimidyl ester (1:20, ThermoFisher) by media change. Immediately after stimulation, cells were returned to the IncuCyte S3 for imaging in the ‘phase’ and ‘red’ channel with 9 images/well every 15 min for 2–3 hr. Images were analysed using the IncuCyte 2019B Rev2 software (Sartorius) as total integrated intensity in ‘red’ channel normalised to confluence in ‘phase’ channel.
Human tissue
Request a detailed protocolUse of human tissue for synaptoneurosome experiments above was reviewed and approved by the Edinburgh Brain Bank ethics committee and the ACCORD medical research ethics committee, AMREC (approval number 15-HV-016; ACCORD is the Academic and Clinical Central Office for Research and Development, a joint office of the University of Edinburgh and NHS Lothian). The Edinburgh Brain Bank is a Medical Research Council funded facility with research ethics committee (REC) approval (11/ES/0022).
Cathepsin probe assays
Request a detailed protocolCathepsin probe assays were carried out as described in Hamilton et al., 2008. Briefly, microglia were isolated and cultured for 3 days as above before cells were lysed in RIPA buffer (60 μL/well, Merck). Next, 25 μL lysate was combined with 175 μL cathepsin probe L (Z-Phe-Arg-AMC, 45 μM, Bachem) or C (H-Gly-Phe-AMC, 56 μM, Bachem) in an assay buffer comprising 150 mM NaCl, 2 mM EDTA, 5 mM DTT, and 100 mM trisodium citrate at pH 5.5 in a black-walled, black-bottom 96-well plate (Corning). Lysates (or lysis buffer alone as a background control) were incubated for 2 hr at 37°C before reading on a fluorescence plate reader (BMG LABTECH) at excitation 360 nm, emission 460 nm with gain adjusted as necessary. Data are presented as background-corrected as relative fluorescence units.
Immunohistochemistry
Request a detailed protocolBrains from 12-month-old male and female AppWt/WtCst7+/+, AppNL-G-FCst7+/+, and AppNL-G-FCst7-/- mice were isolated by terminally anaesthetising with 3% isoflurane (33.3% O2 and 66.6% N2O) and transcardial perfusion with ice-cold DEPC-treated 0.9% NaCl, 0.4% trisodium citrate. Brains were immediately separated down the midline then the left hemisphere placed into ice-cold 10% neutral buffered formalin (ThermoFisher). Hemi-brains were post-fixed for 48 hr at 4°C before processing for paraffin embedding. For fluorescence immunostaining used to investigate microglial and lysosomal burden, 6 μm sagittal sections were deparaffinised with 2×10 min xylene before rehydration in subsequent changes of 100%, 90%, and 70% ethanol (all 5 min). Sections were rinsed in dH2O before antigen retrieval in 10 mM TrisEDTA pH 9 (30 min at 95°C). Next, sections were rinsed and incubated with primary antibody (Table 3) in PBS, 1% BSA (Merck), 0.3% Triton-X (Merck) at 4°C overnight. Sections were washed in PBS, 0.1% Tween-20 (Merck) 3×5 min before incubation with secondary antibodies (Table 3) in PBS, 0.1% Tween-20 (Merck), 1% BSA (Merck) at RT for 1 hr. Sections were washed again, incubated in TrueBlack Lipofuscin Autofluorescence Quencher (1:300 in 70% ethanol, Biotium) for 1 min, washed again in PBS, and rinsed in dH2O before mounting with Fluorescence Mounting Medium (Dako). For synaptophysin staining, Thioflavin S (Merck) was added 1:200 for 5 min before mounting. Slides were imaged using an AxioImager D2 microscope (Zeiss) with a 20× objective. For quantification, two random fields of view were taken per mouse for both cortex and hippocampus. One image was taken for subiculum. One mouse was excluded from analysis as the section taken did not contain hippocampus. To remove the influence of plaque burden on the results, images were taken of approximately equal 6E10 burden. Images were analysed using a threshold analysis with QuPath v0.3.0 (Bankhead et al., 2017) and co-staining was quantified using Definiens Developer. For imaging and quantification of synaptophysin staining, five images were taken of a single plaque/image in AppNL-G-F brains using a 40× oil-immersion objective with five random images in plaque-free wild-type brains. Synaptophysin coverage was calculated using ImageJ auto-threshold (default settings) and intensity within vs. outwith plaque area calculated using QuPath v0.3.0 (Bankhead et al., 2017).
Table 3
Antibodies used for immunohistochemistry.
| Antigen | Type | Species (raised) | Supplier | Cat number | Stock concentration (mg/mL) | Dilution |
|---|---|---|---|---|---|---|
| IBA1 | Primary | Rabbit | Wako | 019-19741 | 0.5 | 1:2000 |
| Aβ (6E10) | Primary | Mouse | BioLegend | 803001 | 1 | 1:1000 |
| LAMP2 | Primary | Rat | BioLegend | 108501 | 0.5 | 1:200 |
| Synaptophysin (Sy38) | Primary | Mouse | Abcam | ab8049 | Lot:1043113-1 | 1:200 |
| Anti-rabbit IgG Alexa Fluor 488 | Secondary | Goat | ThermoFisher | A-11008 | 2 | 1:500 |
| Anti-mouse IgG Alexa Fluor 555 | Secondary | Donkey | ThermoFisher | A-31570 | 2 | 1:500 |
| Anti-rat IgG Alexa Fluor 647 | Secondary | Goat | ThermoFisher | A-21247 | 2 | 1:500 |
| Anti-mouse Biotinylated | Secondary | Goat | Vector Labs | ba-9200 | 1.5 | 1:100 |
For chromogenic 3,3'-diaminobenzidine (DAB) staining used to quantify amyloid burden, 6 μm sagittal sections were deparaffinised and rehydrated as above. Following antigen retrieval, endogenous peroxidase activity was blocked with 0.3% H2O2 for 10 min and sections were blocked with 5% normal goat serum (Vector Labs) in PBS for 1 hr at RT. 6E10 primary antibody was added at 1:500 and incubated overnight at 4°C. The next day, sections were washed and stained with biotinylated anti-mouse IgG (Table 3) in PBST 1% BSA for 1 hr at RT before washing and addition of ABC Elite amplification (Vector Labs) for 30 min at RT according to the manufacturer’s instructions. After a further wash, slides were immersed in DAB solution (0.5 mg/mL DAB, 0.015% H2O2 in PBS) until stain had developed. Sections were counterstained with acidified Harris Hematoxylin (Epredia), dehydrated through increasing concentrations of ethanol and xylene, and mounted with Pertex mountant (CellPath). For imaging, slides were scanned with an Axioscanner Slide Scanner (Zeiss) in brightfield at 20× magnification. Images were analysed using a threshold analysis with QuPath v0.3.0 (Bankhead et al., 2017). DAB slides were stained in two batches with equal representation of groups between batches, data are presented as batch-normalised 6E10 % positive area.
In situ hybridisation
Request a detailed protocolIn situ hybridisation was performed using RNAscope 2.5 HD Duplex Assay using slight modifications to the manufacturer’s protocol. Freshly cut (6 µm) brain tissue was baked onto slides at 60°C for 30 min before deparaffinisation (2× xylene, 5 min) and dehydration (2×100% EtOH, 1 min). Hydrogen peroxide was applied for 10 min at RT and antigen retrieval was performed for 30 min in a pre-heated (20 min) plastic Coplin jar in a 97.5°C waterbath. Sections were left at RT overnight before placing into humidity chambers and protease plus was applied for 15 min at 37°C. Probes targeting Cst7 (498711-C2) and Lilrb4a (1260291-C2) were incubated for 2 hr at 40°C, amplified according to the manufacturer’s instructions, and red signal detected using RNAScope Fast Red. At this stage, antibodies raised against IBA1 and Aβ (6E10) were added as described above and the experiment continued as with fluorescence immunohistochemistry as above. RNAScope Fast Red is detectable both in brightfield and fluorescence in 555 channel. For imaging, slides were either scanned with an Axioscanner Slide Scanner (Zeiss) in brightfield at ×20 magnification or imaged in fluorescence using an AxioImager D2 microscope (Zeiss) with a ×20 objective. For quantification, two random fields of view were taken per mouse cortex. Images were analysed for Cst7/Lilrb4a coverage within/outwith plaque region using QuPath v0.3.0 (Bankhead et al., 2017) to draw around 6E10+ area and co-localisation with IBA1 was analysed using MultipleColourAnalysis ImageJ plugin.
Multiplex ELISA
Request a detailed protocolMouse cytokines from microglial supernatants were measured using MILLIPLEX multiplex assays as described previously (McCulloch et al., 2022). Briefly, MILLIPLEX MAP Mouse Cytokine/Chemokine Magnetic Bead Panel (MCYTOMAG-70K, Merck) was used to measure GM-CSF, IFN-γ, IL-1α, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-12(p40), IL-33, and TNF-α. In all assays, samples were assayed as single replicates and all samples, standards, and quality controls were prepared in accordance with the manufacturer’s instructions. Samples were incubated with beads on a plate for 1 hr (isotyping assay) or overnight at 4°C and washes carried out using a magnetic plate washer. Plates were analysed using a Magpix Luminex machine and Luminex xPonent software version 4.2, with a sample volume of 50 μL per well and a minimum of 50 events counted per sample.
Randomisation and blinding
Request a detailed protocolExperimenters were blinded to genotype groups throughout the study. Animals were given an experimental identifier and all samples were analysed using this coded identifier. Mice were randomly assigned to cull groups using random number generator on Microsoft Excel (which were performed in batches due to throughput for FACS) with stratification for experimental group. Order of MeX04 injection and subsequent mouse termination and tissue collection was randomised by random number generator using Microsoft Excel. Data was unblinded for analysis after experimental work was complete. There were no specific inclusion/exclusion criteria. However, mice were excluded from IHC analysis in Figures 3 and 6 due to attrition (tissue sections were not obtainable of the specific regions required). Power calculations (α=0.05, power = 0.8) based on the primary outcome of 50% effect size on AppNL-G-F vs. AppNL-G-FCst7-/- %MeX04+ microglia demonstrated n=12 would be sufficient for our study.
Statistical analyses
Request a detailed protocolData are presented as mean values + standard error of the mean (SEM). Levels of significance were p<0.05 (*), p<0.01 (**), p<0.001 (***). In all figures, replicates (defined by n) were biological replicates. Statistical analyses were carried out using GraphPad Prism (version 9). For RNASeq data, statistical significance was calculated using DESeq2 R package (Love et al., 2014). Immunohistochemistry, qPCR, cathepsin activity, and cytokine secretion were analysed with a two-way ANOVA followed by Tukey’s or Sidak’s post hoc comparisons, unpaired Student’s t-test, or mixed effects modelling. Live-imaging data were analysed by area under the curve followed by Student’s t-test or one-way ANOVA with Dunnett’s multiple comparisons test. Transformations were applied where necessary. Graphs and figures were created with GraphPad Prism (version 9), VolcaNoseR (Goedhart and Luijsterburg, 2020), DeepVenn (Hulsen, 2022), and BioRender.com.
Data availability
Sequencing data obtained and reanalysed in Figure 1 was obtained from published manuscripts. All data have been deposited in GEO: GSE89482 (Srinivasan et al., 2020), GSE65067 (Wang et al., 2015), GSE74615 (Orre et al., 2014), GSE117646 (Kang et al., 2018), GSE203202 (Guillot-Sestier et al., 2021), GSE127893 (Sala Frigerio et al., 2019). Newly generated data that support the findings of this study are uploaded to ArrayExpress (Accession ID:E-MTAB-13360).
-
ArrayExpressID E-MTAB-13360. RNA-seq to investigate sex-dependent changes in microglia in Cst7 knockout mice crossed with a mouse model of amyloid-driven Alzheimer's Disease.
-
NCBI Gene Expression OmnibusID GSE65067. Expression data from WT and TREM2 deficient microglia in a mouse model of Alzheimer's disease.
-
NCBI Gene Expression OmnibusID GSE89482. Gene Expression in Cx3cr1-GFP+ Cells in PS2APP Alzheimer's Disease mice.
-
NCBI Gene Expression OmnibusID GSE74615. Acutely isolated murine cortical astrocytes and microglia: Alzheimer's disease vs wildtype.
-
NCBI Gene Expression OmnibusID GSE117646. Microglial translational profiling reveals a convergent APOE pathway from aging, amyloid, and tau.
-
NCBI Gene Expression OmnibusID GSE203202. Sex-related differences in Alzheimer's disease.
-
NCBI Gene Expression OmnibusID GSE127893. The major risk factors for Alzheimer's disease: Age, Sex and Genes, modulate the microglia response to Aβ plaques.
References
-
Sex modifies the APOE-related risk of developing Alzheimer diseaseAnnals of Neurology 75:563–573.https://doi.org/10.1002/ana.24135
-
Immortalization of murine microglial cells by a v-raf/v-myc carrying retrovirusJournal of Neuroimmunology 27:229–237.https://doi.org/10.1016/0165-5728(90)90073-v
-
Microglia function in the central nervous system during health and neurodegenerationAnnual Review of Immunology 35:441–468.https://doi.org/10.1146/annurev-immunol-051116-052358
-
The neuropathological diagnosis of Alzheimer’s diseaseMolecular Neurodegeneration 14:32.https://doi.org/10.1186/s13024-019-0333-5
-
Interleukin-18 and IL-18 binding proteinFrontiers in Immunology 4:289.https://doi.org/10.3389/fimmu.2013.00289
-
STAR: ultrafast universal RNA-seq alignerBioinformatics 29:15–21.https://doi.org/10.1093/bioinformatics/bts635
-
Sex-dependent effect of APOE on Alzheimer’s disease and other age-related neurodegenerative disordersDisease Models & Mechanisms 13:dmm045211.https://doi.org/10.1242/dmm.045211
-
Distinct amyloid-β and tau-associated microglia profiles in Alzheimer’s diseaseActa Neuropathologica 141:681–696.https://doi.org/10.1007/s00401-021-02263-w
-
Isolation and phenotyping of adult mouse microglial cellsMethods in Molecular Biology 1784:77–86.https://doi.org/10.1007/978-1-4939-7837-3_7
-
Microglia in Alzheimer’s diseaseThe Journal of Cell Biology 217:459–472.https://doi.org/10.1083/jcb.201709069
-
A role for TREM2 ligands in the phagocytosis of apoptotic neuronal cells by microgliaJournal of Neurochemistry 109:1144–1156.https://doi.org/10.1111/j.1471-4159.2009.06042.x
-
Microglia use TAM receptors to detect and engulf amyloid β plaquesNature Immunology 22:586–594.https://doi.org/10.1038/s41590-021-00913-5
-
Microglial translational profiling reveals a convergent APOE pathway from aging, amyloid, and tauThe Journal of Experimental Medicine 215:2235–2245.https://doi.org/10.1084/jem.20180653
-
TREM2 mutations implicated in neurodegeneration impair cell surface transport and phagocytosisScience Translational Medicine 6:243ra86.https://doi.org/10.1126/scitranslmed.3009093
-
Do microglial sex differences contribute to sex differences in neurodegenerative diseases?Trends in Molecular Medicine 25:741–749.https://doi.org/10.1016/j.molmed.2019.05.001
-
Disinhibition of cathepsin C caused by cystatin F deficiency aggravates the demyelination in a cuprizone modelFrontiers in Molecular Neuroscience 9:152.https://doi.org/10.3389/fnmol.2016.00152
-
Neurotrophic factors regulate cathepsin S in macrophages and microglia: A role in the degradation of myelin basic protein and amyloid beta peptideMolecular Medicine 5:334–343.
-
Exploring sex-related differences in microglia may be a game-changer in precision medicineFrontiers in Aging Neuroscience 14:868448.https://doi.org/10.3389/fnagi.2022.868448
-
Isolation of glia from Alzheimer’s mice reveals inflammation and dysfunctionNeurobiology of Aging 35:2746–2760.https://doi.org/10.1016/j.neurobiolaging.2014.06.004
-
The ARRIVE guidelines 2.0: Updated guidelines for reporting animal researchJournal of Cerebral Blood Flow and Metabolism 40:1769–1777.https://doi.org/10.1177/0271678X20943823
-
Considering sex and gender in Alzheimer disease and other dementiasDialogues in Clinical Neuroscience 18:437–446.https://doi.org/10.31887/DCNS.2016.18.4/cepperson
-
Single App knock-in mouse models of Alzheimer’s diseaseNature Neuroscience 17:661–663.https://doi.org/10.1038/nn.3697
-
Clearance of apoptotic neurons without inflammation by microglial triggering receptor expressed on myeloid cells-2The Journal of Experimental Medicine 201:647–657.https://doi.org/10.1084/jem.20041611
-
Cysteine cathepsins: from structure, function and regulation to new frontiersBiochimica et Biophysica Acta 1824:68–88.https://doi.org/10.1016/j.bbapap.2011.10.002
-
Synaptic degeneration in Alzheimer diseaseNature Reviews. Neurology 19:19–38.https://doi.org/10.1038/s41582-022-00749-z
-
Sex-specific features of microglia from adult miceCell Reports 23:3501–3511.https://doi.org/10.1016/j.celrep.2018.05.048
-
Sexual differentiation of microgliaFrontiers in Neuroendocrinology 52:156–164.https://doi.org/10.1016/j.yfrne.2018.11.003
-
An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortexThe Journal of Neuroscience 34:11929–11947.https://doi.org/10.1523/JNEUROSCI.1860-14.2014
Article and author information
Author details
Michael JD Daniels

Funding
UK Dementia Research Institute
- Barry McColl
- Tara Spires-Jones
Leducq Foundation (Stroke IMPaCT - 19CVD01)
- Barry McColl
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
This work is supported by the UK Dementia Research Institute which receives its funding from UK DRI Ltd, funded by the UK Medical Research Council, Alzheimer’s Society and Alzheimer’s Research UK and the Leducq Foundation Transatlantic Network of Excellence, Stroke IMPaCT (19CVD01). For the purpose of open access, the author has applied a CC-BY public copyright license to any Author Accepted Manuscript version arising from this submission. We would like to thank the QMRI Flow Cytometry & Cell Sorting Facility at The University of Edinburgh for assistance with FACS studies, the Shared University Research Facilities (SuRF) Histology facility at The University of Edinburgh for assistance with processing, sectioning, and slide-scanning brains, Dr Daniel Soong at The MRC Centre for Reproductive Health, University of Edinburgh for his assistance with image analysis, and Prof. Siddharthan Chandran at The University of Edinburgh for providing the SH-SY5Y cell line.
Ethics
Human subjects: Use of human tissue for synaptoneurosome experiments above was reviewed and approved by the Edinburgh Brain Bank ethics committee and the ACCORD medical research ethics committee, AMREC (approval number 15-HV-016; ACCORD is the Academic and Clinical Central Office for Research and Development, a joint office of the University of Edinburgh and NHS Lothian). The Edinburgh Brain Bank is a Medical Research Council funded facility with research ethics committee (REC) approval (11/ES/0022).
All animal experiments were carried out in accordance with the United Kingdom Animals (Scientific Procedures) Act 1986 and approved by the United Kingdom Home Office and the local Animal Ethical Review Group, University of Edinburgh.
Copyright
© 2023, Daniels et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,901
- views
-
- 245
- downloads
-
- 21
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 21
- citations for umbrella DOI https://doi.org/10.7554/eLife.85279
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cystatin F (Cst7) drives sex-dependent changes in microglia in an amyloid-driven model of Alzheimer’s disease
eLife 12:e85279.
https://doi.org/10.7554/eLife.85279




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}