Molecular tuning of sea anemone stinging
- Department of Molecular and Cellular Biology, Harvard University, United States
- Machine Learning Center Genoa (MalGa), Department of Civil, Chemical and Environmental Engineering (DICCA), University of Genoa, Italy
- Department of Organismic and Evolutionary Biology and Museum of Comparative Zoology, Harvard University, United States
eLife assessment
This is an important paper that links distinctive stinging behavior of two related anemones occupying different ecological niches to varying inactivation properties of voltage-gated calcium channels conferred by auxiliary Cavβ subunits. Further convincing evidence is provided that these differences are mediated by alternative splicing of Cavβ subunit of the calcium channel. The study will be of interest to scientists studying Ca2+ signaling, ion channel biophysicists, and marine biologists.
https://doi.org/10.7554/eLife.88900.3.sa0Significance of the findings:
Important: Findings that have theoretical or practical implications beyond a single subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Convincing: Appropriate and validated methodology in line with current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Jellyfish and sea anemones fire single-use, venom-covered barbs to immobilize prey or predators. We previously showed that the anemone Nematostella vectensis uses a specialized voltage-gated calcium (CaV) channel to trigger stinging in response to synergistic prey-derived chemicals and touch (Weir et al., 2020). Here, we use experiments and theory to find that stinging behavior is suited to distinct ecological niches. We find that the burrowing anemone Nematostella uses uniquely strong CaV inactivation for precise control of predatory stinging. In contrast, the related anemone Exaiptasia diaphana inhabits exposed environments to support photosynthetic endosymbionts. Consistent with its niche, Exaiptasia indiscriminately stings for defense and expresses a CaV splice variant that confers weak inactivation. Chimeric analyses reveal that CaVβ subunit adaptations regulate inactivation, suggesting an evolutionary tuning mechanism for stinging behavior. These findings demonstrate how functional specialization of ion channel structure contributes to distinct organismal behavior.
Introduction
Sea anemones, jellyfish, corals, and hydrozoans of the Cnidarian phylum use specialized cells called nematocytes to sting for predation or defense. Mechanical and chemical stimuli from prey or predators act synergistically on nematocytes to mediate rapid discharge of a toxin-covered barb from its nematocyst organelle (Holstein and Tardent, 1984; Watson and Mire-Thibodeaux, 1994; Babonis and Martindale, 2014). Nematocyst discharge requires calcium (Ca2+) influx and, as a one-time use organelle, is tightly controlled to prevent energetically wasteful stinging to irrelevant stimuli (Lubbock et al., 1981; Gitter et al., 1994; Watson and Hessinger, 1994). We previously found that the starlet sea anemone Nematostella vectensis uses a uniquely adapted voltage-gated Ca2+ channel (CaV) to integrate simultaneously presented chemical and mechanical cues that elicit nematocyst discharge. Nematostella CaV exhibits unusually ‘strong’ steady-state voltage-dependent inactivation at resting membrane voltages to reduce cellular excitability and prevent stinging behavior in response to extraneous, non-prey touch signals. Chimeric analyses of Nematostella and mammalian CaV showed that the auxiliary β subunit (CaVβ) is required and sufficient for low-voltage steady-state inactivation in Nematostella CaV channel complexes. CaV inactivation is relieved by hyperpolarization of the nematocyte membrane potential to very negative voltages through the effect of prey-derived chemosensory signals that are synaptically transmitted from sensory neurons. Upon relieving CaV inactivation, direct touch responses are amplified to trigger nematocyst discharge (Weir et al., 2020). Thus, single nematocytes integrate synergistic cues to elicit a precise response, representing a unique cellular system to study how cells detect and transduce signals to produce discrete behavior.
While CaV-mediated sensory integration represents one mechanism by which nematocytes ‘decide’ when to sting, the incredible diversity of cnidarian biology suggests that stinging behavior must be adapted to support the demands of different lifestyles. Cnidarian taxa occupy diverse environmental niches and endure specific metabolic demands, predatory challenges, and environmental pressures for survival, which results in distinct selective pressures on nematocyte evolution (Beckmann and Özbek, 2012; Babonis et al., 2022). Nematostella vectensis and Exaiptasia diaphana represent an example of closely related cnidarians with differing environmental niches and metabolic demands (Darling et al., 2005; Bedgood et al., 2020). Nematostella are found in shallow brackish water of coastal marshes where they are buried in the mud, hidden from predators, with only their tentacles exposed to catch unsuspecting passing prey (Fraune et al., 2016). Thus, we hypothesize that their stinging is under tight regulation adapted for opportunistic predation. In contrast, Exaiptasia are exposed to predators while living in shallow, open ocean environments that provide sufficient sunlight for their endosymbionts to produce important photosynthetic products and nutrients (Baumgarten et al., 2015). Considering these dramatically different ecological contexts, we hypothesized that CaV-mediated regulation of nematocyte discharge has adapted to reflect the demands on stinging behavior in these two anemones. We therefore probed the behavior of these related but distinct anemones and investigated how subtle tuning of a shared molecular-regulatory mechanism drives adaptation in physiology and behavior associated with niche diversification.
In this study, we find that the symbiotic anemone Exaiptasia stings in response to mechanical stimuli alone, independent of predation pressure. This behavior serves as a stark contrast with Nematostella stinging, which is only elicited by synergistic prey chemicals and touch. Markov decision process modeling coupled with behavioral experiments revealed that Nematostella stings as an optimal predator, whereas Exaiptasia exhibits optimal defensive stinging behavior. Consistent with indiscriminate stinging behavior, we discover that Exaiptasia nematocyte physiology lacks the unusual CaV inactivation used by Nematostella to inactivate cells at rest and prevent responses to touch in the absence of prey chemicals. ‘Weak’ steady-state inactivation of Exaiptasia CaV is mediated by a splice isoform of the beta subunit (CaVβ) with a distinct N-terminus and allows for robust activation from resting membrane potentials. Analysis of chimeric jellyfish and anemone channels reveals that CaV inactivation is broadly regulated by the CaVβ N-terminus, suggesting an evolutionary tuning mechanism that could contribute to specific stinging behavior across cnidarians. Thus, we propose CaV adaptations as one molecular mechanism that could shift predatory versus defensive stinging in cnidarians. These results highlight how subtle adaptations in protein structure contribute to complex organismal behavior.
Results
Comparative sea anemone stinging behavior
In their natural habitat, Nematostella are hidden by burrowing within the sandy substrate and use an opportunistic predatory strategy to capture prey with their tentacles. In contrast, symbiotic Exaiptasia are found within open waters where they experience greater risk of predation and therefore must adopt a more defensive stance. Thus, we first asked whether differences in ecological pressure are reflected by stinging behavior. Consistent with our previous findings, we observed Nematostella stinging in response to simultaneously delivered prey extract and touch, reflecting stinging control adapted for predation (Figure 1; Weir et al., 2020). Strikingly, Exaiptasia tentacles instead exhibited robust stinging even in the absence of prey chemicals (touch alone, Figure 1). Similar touch-evoked stinging was observed for Exaiptasia acontia, which are defensive nematocyte-enriched structures that are ejected and release toxins to repel predators (Lam et al., 2017). Considering the drastic differences in stinging behavior, we wondered if Exaiptasia’s indiscriminate stinging reflects a distinct control strategy.
Figure 1
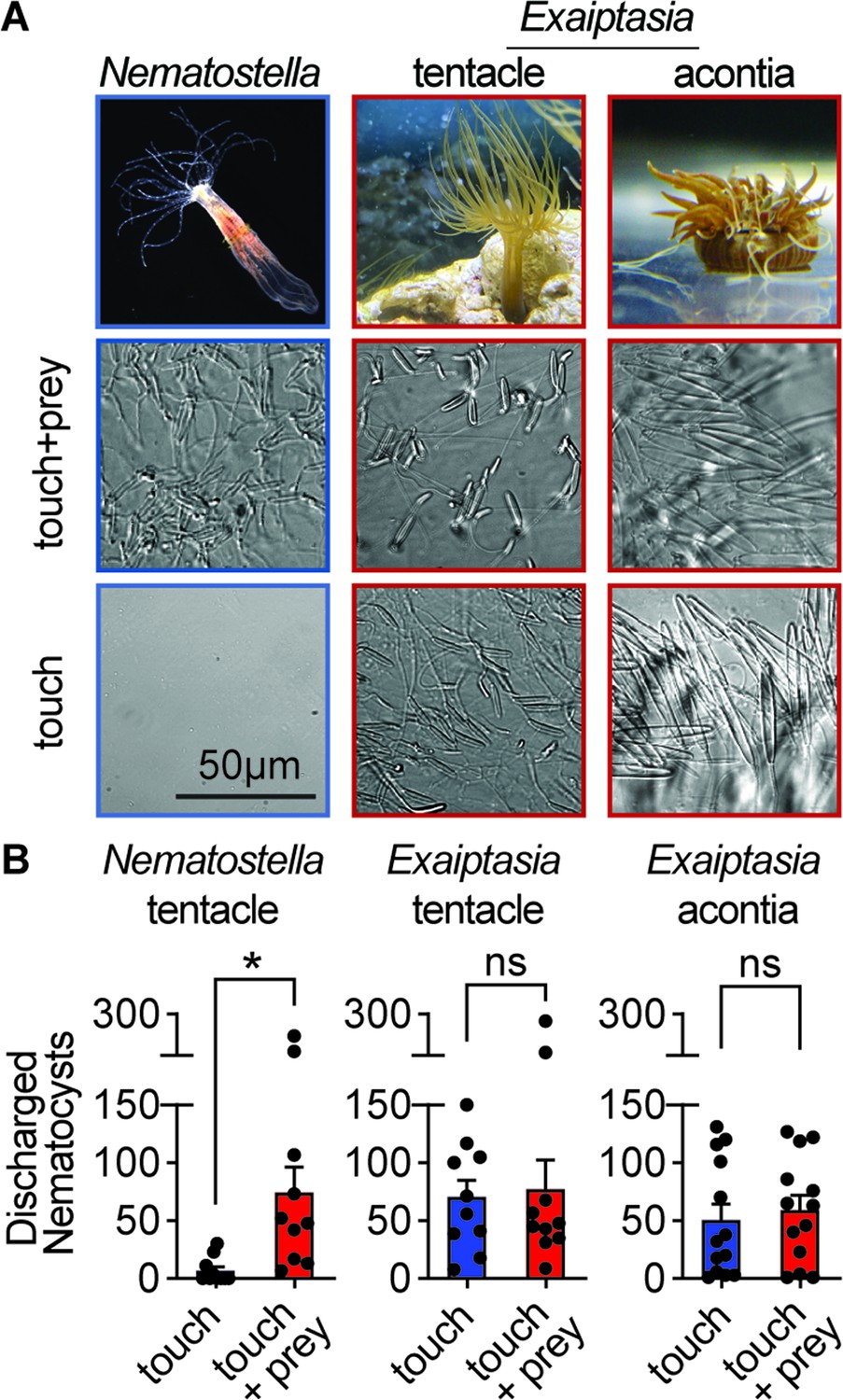
Comparative sea anemone stinging behavior.
(A) Nematostella vectensis stings with tentacles while Exaiptasia diaphana also stings with acontia filaments that are ejected from its body for defense. Left: Nematostella nematocyte discharge was only observed in response to simultaneous prey chemicals and touch stimuli. Middle, Right: Exaiptasia nematocyte discharge from tentacles and acontia occurred irrespective of prey cues (touch alone). Scale bar = 50 μm. (B) Nematostella nematocyte discharge was elicited by simultaneous touch and prey chemical stimuli (n=10 trials). Exaiptasia tentacle (n=10) and acontia (n=13) nematocytes discharged only to touch, with or without prey chemicals. p<0.05 for Nematostella, paired two-tailed student’s t-test. Data represented as mean ± sem.
To investigate whether different stinging behaviors might be suited for predation versus defense, we developed a normative theory aimed at predicting optimal stinging behavior as a function of nutritional state (see Materials and methods for model details). We focused on stinging intensity, defined as the fraction of nematocysts discharged during a stinging event, and asked whether nutritional state would affect optimal predatory and defensive stinging. In the language of decision models, the intensity of stinging is an action, and it is chosen by the agent, or anemone. In our study, ‘choice’ of stinging means modulation of behavior with nutritional state, rather than a cognitive process. Each choice has associated costs and benefits that depend on the environment. Because anemones sting many times over the course of their lives, an optimal behavior must account for overall costs and benefits after many events; therefore, this is a sequential decision-making problem.
We modeled the optimal stinging response to a given environment by using Markov decision processes (MDP). Each anemone was modeled as an ‘agent’ that must hedge the intensity of its stinging response. The environment, including the identity of prey, predators, and the physiological state of the animals, defines the likelihood, costs, and benefits of successful stinging. Specifically, intense stinging responses are costly since each fired nematocyst needs to be regenerated. But they are also more likely to succeed because greater discharge of stinging barbs increases the likelihood of contact and envenomation. The cost per nematocyte was first assumed to be constant and equivalent for defensive and predatory stinging as nematocyst discharge requires regeneration in either case (Figure 2A, solid line, filled circles). We assumed that the benefits of successful predatory stinging depend on the capture and consumption of prey, which improves satiation (Figure 2B left). In contrast, stinging a predator for defense would not improve nutritional state, hence the benefits of stinging would not depend on starvation (Figure 2C left). We then used the model to predict optimal stinging that maximizes the sum of all future benefits while minimizing costs during starvation (i.e. maximizing the value function, see Materials and methods). We then tested the prediction directly against experiments with behaving animals.
Figure 2 with 5 supplements see all
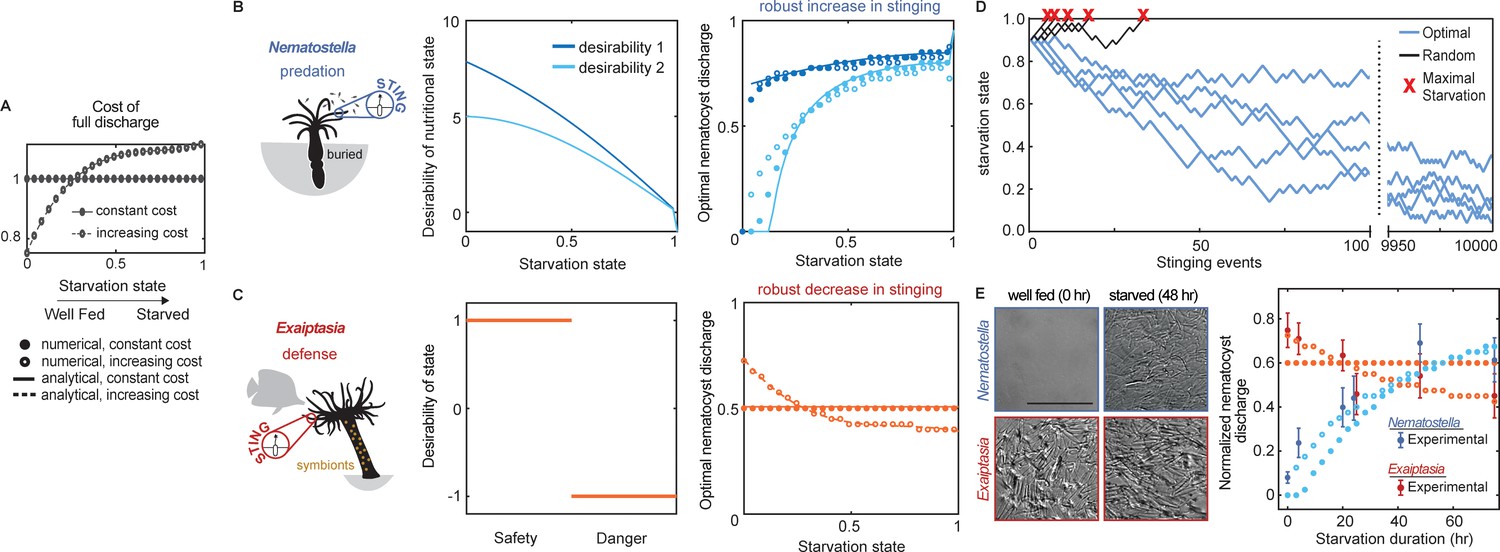
Nematostella stinging is regulated by predation while Exaiptasia stings for defense.
(A) The cost of stinging is , where is the cost for full nematocyte discharge and it either does not change (solid lines filled circles) or increases slightly (dashed lines empty circles) with starvation state. These symbols are used throughout the figure to represent each cost function. The increasing cost is obtained by fitting the Exaiptasia behavior (see fitting procedure in Materials and methods). (B) Left: Nematostella burrows in the substrate and stings for predation. Center: Desirability of nutritional state, or reward, decreases with starvation. Two examples are shown: example 1, ; example 2, . Right: Predicted optimal stinging obtained by solving equation (1) with numerical simulations (circles) and approximate analytical solutions (lines) assuming: and ; with cost for full discharge matching panel A (full circles and solid lines for constant cost; empty circles for increasing cost); reward in Left panels (colors match). For all reward and cost functions, optimal predatory stinging increases with starvation under broad assumptions (see Materials and methods). (C) Left: Exaiptasia diaphana relies heavily on endosymbiotic algae for nutrients and stings primarily for defense. Center: We assumed there are two states, safety (L), and danger (D). The state of safety can transition to danger, but not the other way around. We assumed the agent obtains reward 1 in state L and penalty –1 in state D. Right: Predicted optimal stinging obtained by solving equation (2) with numerical simulations (circles) and analytical solutions (lines). Styles match the costs in panel A; we assume and as before. Optimal defensive stinging is constant or decreases with starvation under broad assumptions (see Materials and methods). (D) Examples of optimal (blue) versus random (black) predatory stinging. Each agent (anemone) starts with , and stings sequentially for many events (represented on the x axis). The random agent almost always reaches maximal starvation before 50 events (grey lines, five examples shown). In comparison, the optimal agent effectively never starves due to a successful stinging strategy optimized for predation (blue lines, five examples shown, parameters as in panel B, curve with matching color). (E) Left: Nematostella nematocyte discharge was affected by prey availability while Exaiptasia stung at a similar rate regardless of feeding. p<0.0001 for Nematostella, two-way ANOVA with post hoc Bonferroni test (n=10 animals, data represented as mean ± sem). Right: Experimental data (circles with error bars representing standard deviation) are well fit by normalized optimal nematocyst discharge predicted from MDP models for both Exaiptasia (orange full and empty circles for constant and increasing cost, panel A) and Nematostella (light blue full and empty circles for constant and increasing cost, panel A and desirability 2 in panel B). We match the last experimental data point to , the precise value of this parameter is irrelevant as long as it is smaller than 1, representing that animals are not severely starved during the experiment.
Using this approach, we found that optimal stinging strategies were completely different for predatory versus defensive behavior. Regardless of the specific environment (likelihood to succeed and specific costs and benefits), predatory stinging increased with starvation (Figure 2B right, solid lines, filled circles). To test our theory regarding predatory stinging, we carried out simulations in which agents discharged a random fraction of nematocytes between 0 and 1, regardless of starvation. Random stinging was unsustainable over numerous events and agents quickly reached maximal starvation state. Agents using optimal predatory stinging discharged more nematocysts when starved and less when satiated, leading to sustained stinging behavior and survival. This was true even if they fired the same fraction of nematocytes as the random agent (Figure 2D). In contrast, optimal stinging for defense stayed constant with starvation (Figure 2C right, solid lines, filled circles). Importantly, while the precise optimal response depended on the details of cost and reward that defined the MDP, the differences between increasing predatory stinging versus unchanging defensive stinging were consistent and largely independent of assumptions associated with each reward function (described in Materials and methods, Figure 2—figure supplements 1–4). These results reflect greater rewards to predatory anemones upon stinging during starvation, whereas defensive anemones sting at a similar rate regardless of nutritional status. Thus, our model predicts robust differences in predatory versus defensive stinging behavior.
We next sought to experimentally test whether pressure to predate regulates stinging in Nematostella and Exaiptasia. To do so, we fed both species of anemones copious amounts of prey (brine shrimp, Artemia nauplii) for 1–2 weeks and then deprived them of food for 5 days. Following manipulation of prey availability, Nematostella significantly increased stinging in response to starvation, while Exaiptasia stinging remained relatively constant despite complete deprivation of prey (Figure 2E, symbols with error bars). The behavior was remarkably consistent with our normative theory of optimal stinging strategies for predation versus defense (Figure 2E, filled circles). Furthermore, changes in Nematostella and Exaiptasia stinging were not due to changes in the abundance of nematocytes because tentacles from both animals were abundantly armed with nematocytes across feeding conditions (Figure 2—figure supplement 5). The experimental behavior of Exaiptasia showed a slight decrease in stinging with starvation. To account for this decrease we revisited the theory and assumed that the cost per nematocyte slightly increased with starvation (Figure 2A, dashed lines and open circles). In this case, the optimal response slightly decreased for defensive stinging but increased for predatory stinging (Figure 2B right, open circles and Figure 2C right, dashed lines, open circles). In fact, the fit between theory and data for both Nematostella and Exaiptasia improved when the cost increased slightly with starvation (Figure 2E, open circles). A more dramatic and less realistic increase of the cost with starvation may lead to a decrease in predatory stinging (Figure 2—figure supplement 4). Thus, we conclude that Nematostella controls stinging for opportunistic predation while Exaiptasia stinging is indiscriminate and serves a greater defensive role for this symbiotic anemone.
Sea anemones with different stinging behavior use distinct CaV channels
We next probed the physiological basis underlying these significantly different stinging behaviors. We previously found that Nematostella stinging is triggered by a specialized CaV channel that exhibits strong inactivation at negative voltages to prevent responses to extraneous non-prey mechanical stimuli (Weir et al., 2020). Ca2+ influx triggers an increase in hydrostatic pressure inside the nematocyst capsule that forces the stinging thread to evert explosively at an acceleration of up to 5.41×106 g, placing it among the fastest biological processes in existence (Lubbock and Amos, 1981; Lubbock et al., 1981; Holstein and Tardent, 1984; Weber, 1990; Gitter et al., 1994; Tardent, 1995; Nüchter et al., 2006). Similar to Nematostella, Exaiptasia stinging required extracellular Ca2+ and was abolished by Cd2+, a CaV channel blocker (Figure 3A). Consistent with a Ca2+-dependent stinging mechanism, whole-cell patch clamp recordings from nematocytes revealed the presence of voltage-gated inward currents that were blocked by Cd2+, suggesting that Exaiptasia nematocytes also use CaV channels to control stinging (Figure 3B). Indeed, CaV currents in Exaiptasia nematocytes exhibited similar voltage-dependent activation properties compared with Nematostella nematocytes (Figure 3C). Thus, in agreement with previous findings, we conclude that Ca2+ influx via CaV channels is broadly important for stinging.
Figure 3
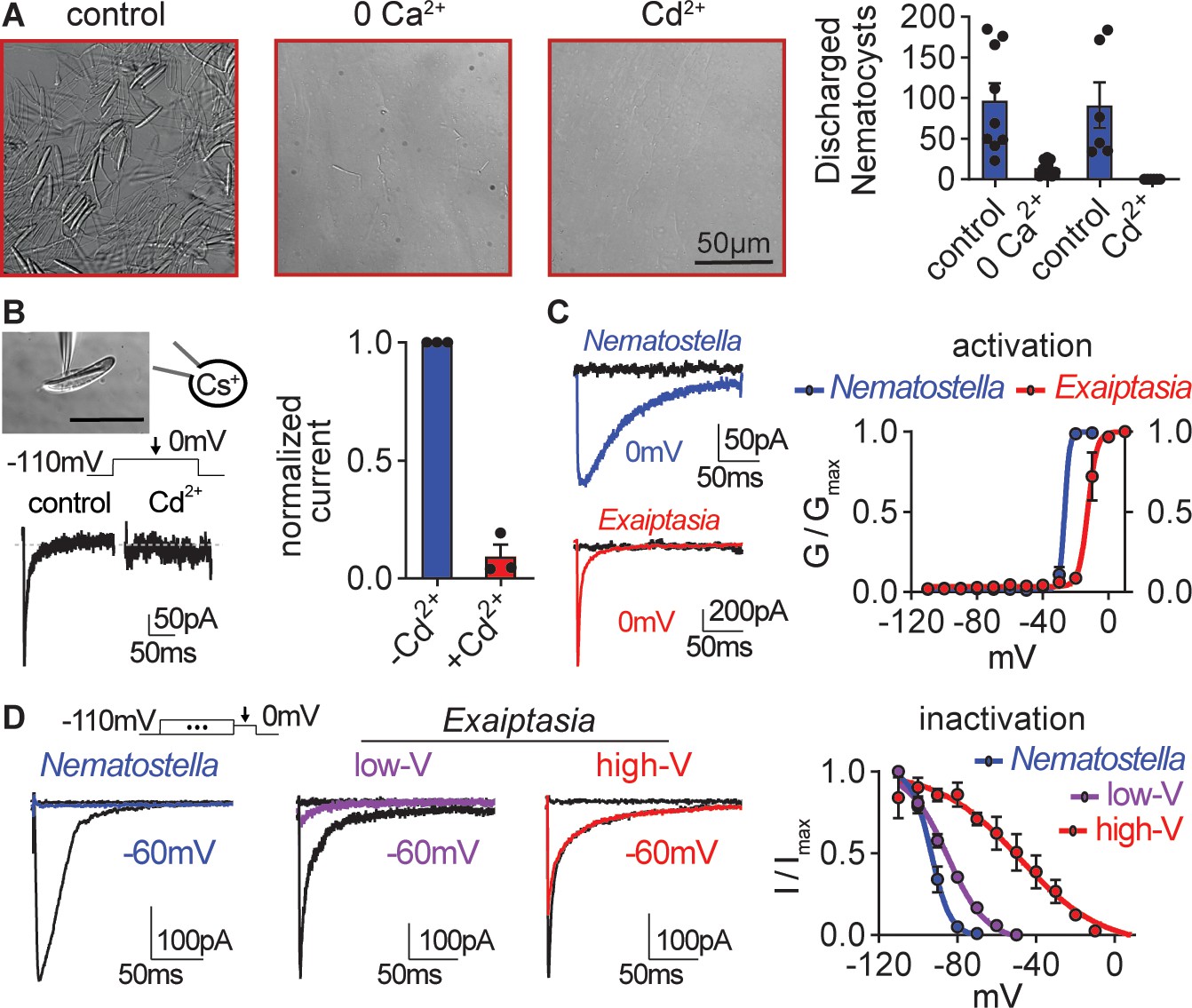
Exaiptasia nematocyte voltage-gated Ca2+ currents exhibit minimal steady-state inactivation compared with Nematostella.
(A) Touch-elicited Exaiptasia tentacle nematocyte discharge was blocked in the absence of Ca2+ (p<0.01, paired two-tailed student’s t-test, n=9 animals) or by addition of the CaV channel blocker Cd2+ (500 μM, p<0.05, paired two-tailed student’s t-test, n=6 animals). Scale bar = 50 μm. (B) Top: Representative patch clamp experiment from an Exaiptasia nematocyte. Scale bar = 20 μm. Bottom: Nematocyte voltage-gated currents elicited by a maximally activating 0 mV pulse were blocked by Cd2+ (n=3 cells, p<0.01, paired two-tailed student’s t-test). (C) Nematocyte voltage-gated currents elicited by –120 mV (black) or 0 mV pulses (colored). Conductance-voltage curves for Nematostella nematocyte (Va1/2 = -26.54 ± 0.78mV, n=3) and Exaiptasia nematocyte (Va1/2 = -12.47 ± 0.70mV, n=3). (D) Nematocyte voltage-gated currents elicited by a maximally activating voltage pulse following 1 s pre-pulses to −110 mV (max current, black), −50 mV (colored), or 20 mV (inactivated, no current). Nematostella nematocytes inactivated at very negative voltages (Vi1/2 = -93.22 ± 0.42mV, n=7) while Exaiptasia contained two populations of nematocytes: low-voltage threshold (Vi1/2 = -84.94 ± 0.70mV, n=4), and high-voltage threshold (Vi1/2 = -48.17 ± 3.32mV, n=3). Data represented as mean ± sem.
CaV channels respond to positive membrane potentials by opening to conduct Ca2+. However, sustained positive voltage drives CaVs to transition to a non-conducting state (inactivation) that prevents re-activation until channels return to a resting state induced by negative membrane potentials. In most cells, voltage-gated ion channel inactivation prevents extended responses to repetitive or prolonged stimulation. Nematostella CaV, is unusual because it inactivates at very negative voltages to prevent responses from resting potential, resulting in nematocytes that cannot fire from rest (Weir et al., 2020). In contrast to Nematostella nematocytes in which half of all CaV channels (Vi1/2) were inactivated at ~ –93 mV, Exaiptasia nematocytes exhibited two distinct inactivation phenotypes: (1) nematocytes with low-voltage threshold (low-V) inactivation similar to that of Nematostella (low-V, Vi1/2 = ~ –85 mV); and (2) a distinct population with weak, high-voltage (high-V) threshold inactivation similar to its well-characterized mammalian orthologue (high-V, Vi1/2 = ~ –48 mV) (Figure 3D). While we did not observe a correlation with abundance or distinct cellular morphology (Östman, 2000; Kass-Simon and Scappaticci, Jr., 2002; Grajales and Rodríguez, 2014), we could clearly distinguish the two populations based on these electrophysiological features. Importantly, high-V nematocyte inactivation was minimal at resting voltages (~ –70 mV), so nearly all channels would be available to amplify depolarizing signals, such as those elicited by touch. Thus, these markedly different physiological properties correlate with distinct stinging behavior: Nematostella uses unusual low-voltage CaV inactivation to integrate sensory cues for tightly regulated predatory stinging. In contrast, Exaiptasia employs a population of nematocytes with weak CaV inactivation, consistent with direct activation from resting potentials and stinging to touch alone.
What is the molecular basis of distinct nematocyte physiology? CaV channels are made of at least three subunits: the pore-forming α and auxiliary β and α2δ subunits. Transcriptomics revealed that nematocyte-enriched tentacles of Exaiptasia expressed cacna1a, the pore-forming subunit homologous to that of the previously characterized Nematostella nematocyte CaV channel (Figure 4—figure supplement 1). We also analyzed the CaV β subunit, CaVβ, which is required and sufficient for the unusual inactivation properties observed in Nematostella CaV (Weir et al., 2020; Figure 4—figure supplement 1). From Exaiptasia, we identified two isoforms of CaVβ: EdCaVβ1 and EdCaVβ2. Droplet digital PCR assays of mRNA abundance showed that both isoforms are expressed throughout Exaiptasia tissues, suggesting they could both be functionally important (Figure 4—figure supplement 1B). To localize CaVβ, we used in situ hybridization to determine that distinct nematocyte populations expressed either EdCaVβ1 or EdCaVβ2 mRNA (but not both) in the same cell (Figure 4B).
Figure 4 with 1 supplement see all
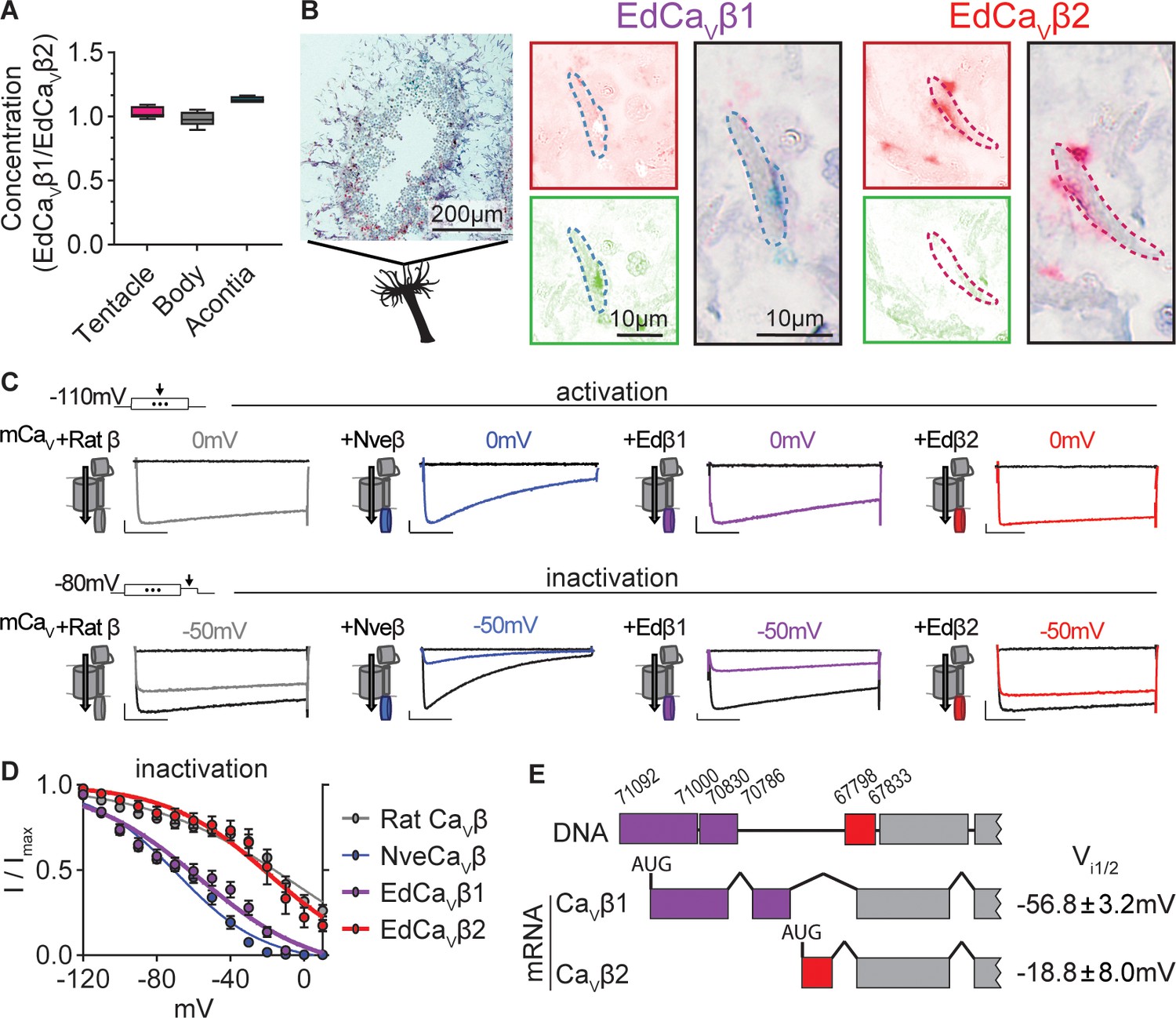
Exaiptasia expresses a CaV β subunit splice isoform that confers weak voltage-dependent inactivation.
(A) ddPCR ratio of concentrations of CaV β subunit 1 and 2 mRNAs was similar in tentacle (n=5), body (n=5), and acontia (n=4 animals) tissue samples. (B) EdCaVβ1 and EdCaVβ2 localized to distinct nematocytes in Exaiptasia tentacle cross section, as visualized by BaseScope in situ hybridization. Representative nematocyte expressing EdCaVβ1 (green) or EdCaVβ2 (red). Representative of three animals. (C) Voltage-gated currents from heterologously-expressed chimeric mammalian CaV (mCaV) with different β subunits: rat (Rattus norvegicus), Nematostella (Nve), Exaiptasia EdCaVβ1 or EdCaVβ2. Top: Currents elicited by voltage pulses to –120 mV (no current, black) and maximally activating 0 mV (colored). Bottom: Voltage-gated currents elicited by a maximally activating voltage pulse following 1 s pre-pulses to −110 mV (max current, black), −50 mV (colored), or 20 mV (inactivated, no current, black). Scale bars = 100 pA, 50ms. (D) Exaiptasia CaV β subunit splice isoforms confer distinct inactivation: Nematostella β subunit (Vi1/2 = -68.93 ± 1.53mV, n=5) and Rat β2a subunit (Vi1/2 = -2.98 ± 13.51mV, n=12) and EdCaVβ1 (Vi1/2 = -56.76 ± 3.18mV, n=8), and EdCaVβ2 (Vi1/2 = -18.84 ± 8.00mV, n=5 cells). Data represented as mean ± sem. (E) Genomic alignment of Exaiptasia β subunit isoforms showed that alternative splicing of the N-terminus region was associated with distinct inactivation: CaVβ1 (long N-term) had low-voltage steady-state inactivation similar to Nematostella, while CaVβ2 (short N-term) exhibited more depolarized steady-state inactivation, matching its mammalian orthologue. Genomic loci listed above sequence.
Considering this expression profile, we wondered if the two CaVβ isoforms could mediate low-V and high-V inactivation phenotypes in Exaiptasia nematocytes. To investigate this question, we heterologously expressed each β subunit isoform with other well-characterized CaV subunits (mammalian CaV α and α2δ) that express well in heterologous systems. Both channels exhibited functional CaV currents with similar activation thresholds (Figure 4C). However, EdCaVβ1- and EdCaVβ2-containing channels significantly differed in their inactivation properties. EdCaVβ1 inactivated at negative voltages, similar to channels containing Nematostella CaVβ (NveCaVβ). In contrast, EdCaVβ2 mediated CaV currents with weak inactivation, more like channels containing rat CaVβ2a (Figure 4C and D). Thus, EdCaVβ1 and EdCaVβ2 confer strong, low-voltage and weak, high-voltage steady-state inactivation, respectively, and are expressed in distinct nematocytes, consistent with low-V and high-V threshold inactivating nematocyte populations. Genomic alignment revealed that alternative splicing at the N-terminus gives rise to EdCaVβ1 and EdCaVβ2 isoforms, serving as a mechanism to dynamically tune nematocyte physiology and potentially stinging behavior in contrast to adaptation through gene duplication and divergence (Figure 4E). Furthermore, by expressing two functional variants, Exaiptasia could use distinct nematocyte populations for different behaviors, including a less pronounced role for predation.
Structural adaptations across cnidarian CaV channels
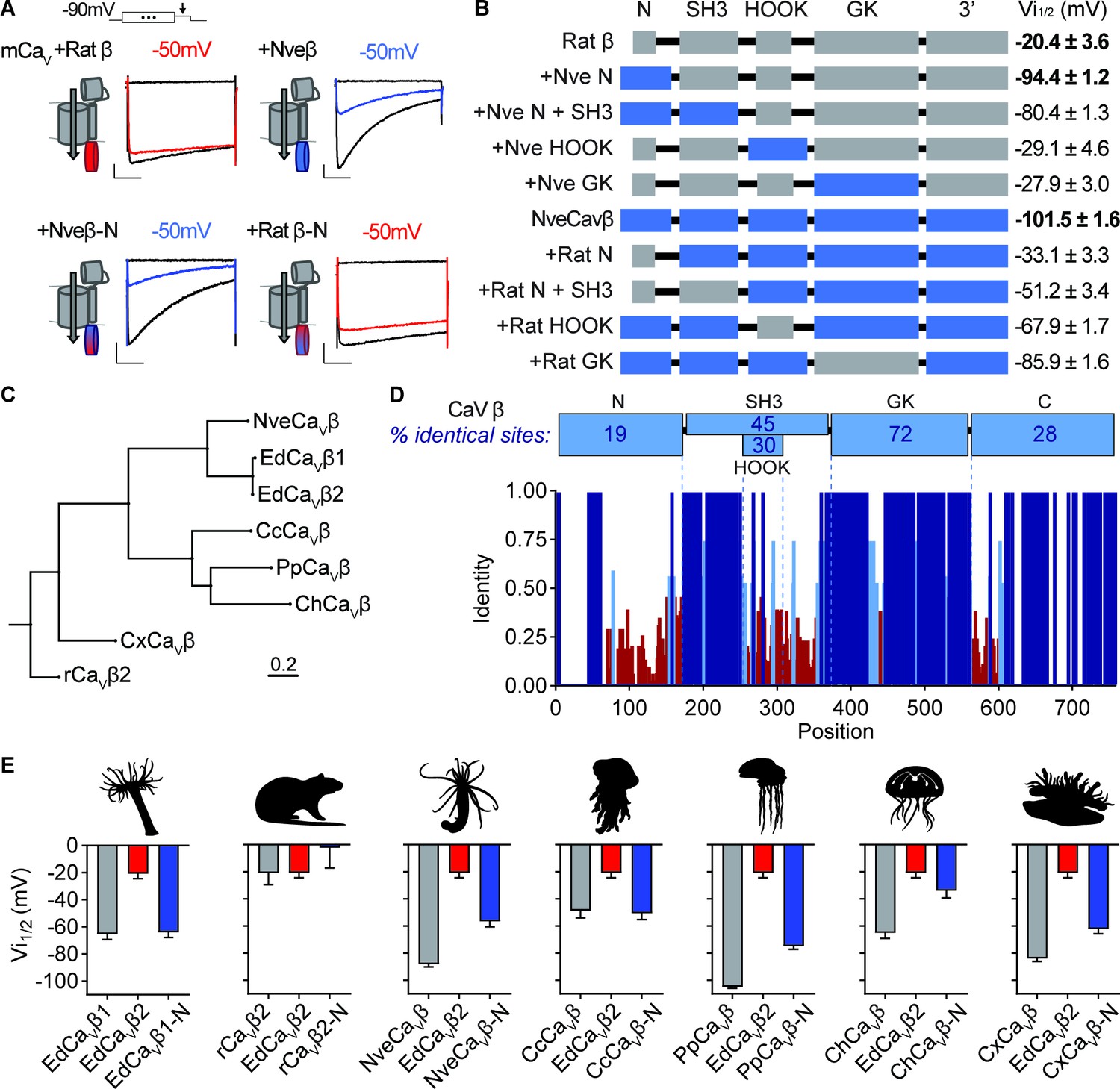
We next asked how variation in CaVβ structure mediates strong phenotypes by testing whether distinct protein domains confer low or high voltage-dependent inactivation. We first compared rat rCavβ2a and Nematostella NveCaVβ, which have significantly different voltage-dependent properties (Weir et al., 2020). Swapping the well-characterized SH3, HOOK, and GK domains had no effect on inactivation, but the NveCaVβ N-terminus was both required and sufficient for low voltage-dependent inactivation (Figure 5A and B). Indeed, swapping only the N-terminus of NveCaVβ was sufficient to shift rat rCavβ2a-conferred inactivation by ~ –75 mV (Figure 5A and B). This finding is consistent with the variation in EdCaVβ splice isoforms, in which differences in the N-terminus account for a~40 mV difference in inactivation thresholds.
Figure 5 with 1 supplement see all
Cnidarian CaV β subunit N-termini confer unique inactivation properties.
(A) Voltage-gated currents from heterologously expressed CaV channels with Nematostella-rat chimeric β subunits demonstrate that the Nematostella N-terminus is sufficient to drive inactivation at negative voltages. Currents shown in response to 10 mV voltage pulses following 1 s pre-pulses to −130 mV (max current, black), −50 mV (colored), or 0 mV (inactivated, no current, black). Scale bars = 100 pA, 50ms. (B) Diagram of CaV Nematostella-rat β subunit domain swaps and resulting Vi1/2 values. The Nematostella β subunit N-terminus is required and sufficient for uniquely hyperpolarized CaV inactivation properties (p<0.001 for average Vi1/2 values across mutant beta subunits, one-way ANOVA with post-hoc Tukey test, n=2–8 cells). (C) Phylogenetic tree of β subunit sequences obtained from several species of cnidarians. Abbreviations of species: Nve, Nematostella vectensis; Ed, Exaiptasia diaphana; Cc, Cyanea capillata (jellyfish); Pp, Physalia physalis (siphonophore); Ch, Clytia hemisphaerica (jellyfish); Cx, Cassiopea xamachana (jellyfish); r, Rattus norvegicus. (D) Top: Percentage of identity between amino acid sequences across β subunit protein domains for NveCaVβ, EdCaVβ1, EdCaVβ2, CcCaVβ, PpCaVβ, ChCaVβ, CxCaVβ2, rCaVβ2. Bottom: Fraction of identity of amino acids across sites of the β subunit protein. (E) Cnidarian CaV β N-termini shift depolarized, weak voltage-dependent inactivation of CaV channels containing EdCaVβ2 to more negative voltages. Voltage-dependent inactivation (Vi1/2) of heterologously-expressed CaVs with WT EdCaVβ2, β subunits from the indicated cnidarians, and chimeras with their N-termini on EdCaVβ2 (p<0.0001 for average Vi1/2 values with multiple comparisons against WT EdCaVβ2 mean, one-way ANOVA with Bartlett’s test and post-hoc Tukey test, n=4–9 cells). Data represented as mean ± sem.
-
Figure 5—source data 1
Amino acid sequences of beta subunits.
- https://cdn.elifesciences.org/articles/88900/elife-88900-fig5-data1-v1.docx
To explore evolutionary relationships of CaVβ, we constructed a phylogenetic tree of sequences from various cnidarians including Nematostella vectensis (anemone, NveCaVβ), Exaiptasia diaphana (anemone, EdCaVβ1 and EdCaVβ2), Cyanea capillata (jellyfish, CcCaVβ), Physalia physalis (hydrozoan, PpCaVβ), Clytia hemisphaerica (jellyfish, ChCaVβ), Cassiopea xamachana (jellyfish, CxCaVβ), and the Rat β subunit (rCavβ2a) as an outgroup (Figure 5C). Sequence comparison across all amino acid positions revealed that the N-terminus exhibited the greatest sequence diversity (Figure 5D), consistent with previous findings showing extensive alternative splicing in this region in other organisms (Helton and Horne, 2002; Helton et al., 2002; Takahashi et al., 2003; Foell et al., 2004; Vendel et al., 2006; Ebert et al., 2008; Buraei and Yang, 2010; Siller et al., 2022). We found that all cnidarian CaVβs conferred voltage-gated currents when co-expressed with CaVα and α2δ subunits and had relatively low voltage thresholds for inactivation compared with rCavβ2a or EdCaVβ2 (Figure 5E, Figure 5—figure supplement 1A, B). Importantly, swapping the N-termini of each cnidarian CaVβ onto EdCaVβ2 was sufficient to shift voltage-dependent inactivation to more negative values (Figure 5E, Figure 5—figure supplement 1C). Thus, alternative splicing at the N-terminus could serve as a broad molecular mechanism for tuning CaV function. Collectively, these findings substantiate the importance of CaVβ in modulating inactivation and suggest a mechanism that could dynamically regulate a small region of only one subunit in the CaV protein complex to tune complex stinging behavior.
Discussion
Collectively, our studies on cnidarian stinging, here and (Weir et al., 2020), reveal different behavior in the primarily predatory anemone Nematostella versus the symbiotic anemone Exaiptasia. This study used a combination of theory and experimentation to uncover the molecular basis of regulation of the divergent behavior of Exaiptasia that uses stinging primarily for defense. Indeed, Exaiptasia obtains a large fraction of its energy and nutrients from endosymbiotic algae (Muscatine et al., 1981; Shick and Dykens, 1984; Steen, 1988), thus reducing overall pressure to predate. This finding is consistent with a common ecological theme in which symbiotic relationships are established whereby one partner provides food and the other provides shelter and defense (Lehnert et al., 2012; Bucher et al., 2016). Therefore, it is plausible that synergistic selection drives higher investment in defensive structures to protect symbiotic species.
Our results demonstrate that molecular adaptations tune distinct stinging behavior: Nematostella CaVβ confers an unusually low threshold for inactivation, basally inhibiting nematocytes unless they are exposed to synergistic prey cues: chemical (hyperpolarizing to relieve inactivation) and mechanical (depolarizing to recruit available CaV channels and elicit stinging; Weir et al., 2020). These physiological mechanisms reflect a stinging strategy suited to opportunistic predation by Nematostella, which burrow within shallow marshes and sting unsuspecting prey. Consistent with the predictions of optimal control theory, Nematostella increased stinging with starvation, suggesting that evolution has shaped its stinging response to maximize benefits for predation. In contrast, Exaiptasia nematocytes contain a functionally specialized splice variant of CaVβ to mediate high threshold voltage-dependent CaV inactivation, consistent with CaV channel availability to amplify depolarizing signals from rest and stinging in response to touch alone. Thus, Exaiptasia physiology is consistent with an indiscriminate stinging strategy for defense, necessary for survival in an exposed environment that facilitates endosymbiotic photosynthesis (Muscatine et al., 1981; Shick and Dykens, 1984; Steen, 1988). Such stinging behavior is likely synergistic with physical escape for some cnidarians (Pallasdies et al., 2019; Wang et al., 2023). Consistent with the predictions of optimal control theory, Exaiptasia stinging was nearly independent of starvation, suggesting that evolution has shaped the stinging response to maximize benefits for defense. Using molecular information gleaned from analyzing these two cnidarians, we find that CaVβ variation across cnidarians mediates differences in voltage-dependent inactivation, which could contribute to differences in stinging behavior. Thus, our study provides an example by which alternative splicing could account for adaptation across this diverse plethora of organisms and habitats.
While theory predicts robust trends for optimal predation and defense independent of environment, the precise nature of the predicted behavior does depend on the environment. In vivo, stinging is likely influenced by stimulus identity and intensity, background turbulence, and other factors. For example, cnidarians may have evolved distinct innate responses for different prey and use chemical sensing to enact the appropriate stinging response. In this case, optimal control theory can be used to predict the optimal response to known salient environmental cues. Alternatively, cnidarians may learn that specific prey are palatable and easy to catch through repeated exposure (Botton-Amiot et al., 2023). In this example, optimal control theory must be replaced by reinforcement learning as the likelihood of successful predation and its cost and benefits (the environment) are unknown (Sutton, 2018).
Indeed, stinging is a complex process mediated by numerous molecular components and cell types that could be subject to evolutionary change or acute modulation. Cnidarians occupy diverse ecological niches and experience varying metabolic demands, predatory challenges, and other survival pressures that could influence stinging behavior. Beyond cnidarians with stationary lifestyles that support photosynthetic endosymbionts (symbiotic anemones, corals, sea pens) and those that use opportunistic ‘sit-and-wait’ ambush predatory strategies (burrowing anemones, siphonophores), others have evolved mobile lifestyles to actively capture prey (jellyfish) (Muscatine et al., 1981; Shick and Dykens, 1984; Steen, 1988; Fraune et al., 2016; Damian-Serrano et al., 2022). Stinging by mobile cnidarians could be subject to different physical demands, such as mechanical disturbance from increased turbulence that could necessitate distinct molecular control. Furthermore, stinging can be influenced by acute factors such as physiological state and various sensory cues, including chemicals, touch, or light (Pantin, 1942; Giebel et al., 1988; Thorington and Hessinger, 1988; Watson and Hessinger, 1989; Plachetzki et al., 2012; Ozment et al., 2021; Aguilar-Camacho et al., 2023). Thus, further inquiry into modulation of stinging across physiological states such as nutritional condition, altered symbiotic relationships, or developmental stages (Sandberg et al., 1971; Columbus-Shenkar et al., 2018) could reveal dynamic regulation by synaptic connections, hormones, or modulation of transcriptional or translational programs (Westfall et al., 1998; Westfall et al., 2002; Westfall, 2004; Weir et al., 2020). Importantly, across all these scenarios, nematocytes remain single-use cells, so it is essential that signaling cascades control discharge in response to the most salient environmental stimuli.
As an early-branching metazoan lineage and sister group to Bilateria (Cartwright et al., 2007), cnidarians are a useful model for probing origins of the nervous system and behavioral specialization (Steele et al., 2011; Jékely et al., 2015; Pallasdies et al., 2019). Here, we present a comparative approach across related cnidarians with distinct physiology and ecology to suggest that behavioral complexity emerges from subtle tuning of single proteins, even in non-neuronal cells. Indeed, cnidarians pose a unique opportunity for the integrative exploration of the evolution of animal behavior. Even beyond neural computations, the emergence of novel cell types among diverse cnidarian body plans, sophisticated predator-prey interactions, and symbioses all contribute to biological novelty and niche expansion (Technau and Steele, 2012). Overall, this work demonstrates how studying evolutionary novelties like stinging behavior can yield broad insight into signal transduction, cellular decision making, and suggests that the evolution of behavior should be examined across all tiers of biological organization.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Nematostella vectensis, adult, male and female) | Nematostella | Woods Hole Marine Biological Laboratory | ||
| Strain, strain background (Exaiptasia diaphana, adult, male and female) | Exaiptasia | Carolina Biological Supply Company | Item #162865 | https://www.carolina.com/marine-and-saltwater-animals/sea-anemone-aiptasia-living/162865.pr |
| Strain, strain background (Cassiopea spp., adult, male and female) | Cassiopea xamachana | Carolina Biological Supply Company | Item #162936 | https://www.carolina.com/marine-and-saltwater-animals/mangrove-jellyfish-living-pack-of-5/162936.pr?question=mangrove |
| Cell line (Homo-sapiens) | HEK293T | ATCC | CRL-3216 | |
| Recombinant DNA reagent | nCaV: cacnb2 (plasmid) | Weir et al., 2020 | NveCaVβ | |
| Recombinant DNA reagent | Rat cacna2d1 | Lin et al., 2004 | Addgene Plasmid #26575 | rCaVα2δ |
| Recombinant DNA reagent | Mouse cacna1a | Richards et al., 2007 | Addgene Plasmid #26578 | mCaVα |
| Recombinant DNA reagent | Rat cacnb2a | Wyatt et al., 1998 | Addgene Plasmid #107424 | rCaVβ |
| Recombinant DNA reagent | Exaiptasia diaphana cacnb1 (plasmid) | This paper | EdCaVβ1 | |
| Recombinant DNA reagent | Exaiptasia diaphana cacnb2 (plasmid) | This paper | EdCaVβ2 | |
| Recombinant DNA reagent | Physalia physalis cacnb (plasmid) | This paper | PpCaVβ | |
| Recombinant DNA reagent | Cyanea capillata cacnb (plasmid) | This paper | CcCaVβ | |
| Recombinant DNA reagent | Cassiopea xamachana cacnb (plasmid) | This paper | CxCaVβ | |
| Recombinant DNA reagent | Clytia hemisphaerica cacnb (plasmid) | This paper | ChCaVβ | |
| Recombinant DNA reagent | EdCaVβ2 with NveCaVβ NTerm (plasmid) | This paper | NveCaVβ-N | |
| Recombinant DNA reagent | EdCaVβ2 with rCaVβ NTerm (plasmid) | This paper | rCaVβ-N | |
| Recombinant DNA reagent | EdCaVβ2 with EdCaVβ1 NTerm (plasmid) | This paper | EdCaVβ1-N | |
| Recombinant DNA reagent | EdCaVβ2 with PpCaVβ NTerm (plasmid) | This paper | PpCaVβ-N | |
| Recombinant DNA reagent | EdCaVβ2 with CcCaVβ NTerm (plasmid) | This paper | CcCaVβ-N | |
| Recombinant DNA reagent | EdCaVβ2 with CxCaVβ NTerm (plasmid) | This paper | CxCaVβ-N | |
| Recombinant DNA reagent | EdCaVβ2 with ChCaVβ NTerm (plasmid) | This paper | ChCaVβ-N | |
| Recombinant DNA reagent | NveCaVβ with Mouse CaVβ HOOK domain (plasmid) | This paper | NVE_cacnb2_mus_hook | |
| Recombinant DNA reagent | Mouse CaVβ with NveCaVβ HOOK domain (plasmid) | This paper | Mus_cacnb2_NVE_hook | |
| Recombinant DNA reagent | NveCaVβ with Mouse CaVβ GK domain (plasmid) | This paper | NVE_cacnb2_mus_GK_domain | |
| Recombinant DNA reagent | Mouse CaVβ with NveCaVβ GK domain (plasmid) | This paper | Mus_cacnb2_NVE_GK_domain | |
| Recombinant DNA reagent | NveCaVβ with rCaVβ NTerm domain (plasmid) | This paper | Rat_Nterm_NVE_cacnb2a | |
| Recombinant DNA reagent | NveCaVβ with rCaVβ NTerm and SH3 domains (plasmid) | This paper | Rat_Nterm_SH3_NVE_cacnb2a | |
| Recombinant DNA reagent | rCaVβ with NveCaVβ NTerm and SH3 domains domain (plasmid) | This paper | NVE_Nterm_SH3_rat_cacnb2a | |
| Recombinant DNA reagent | rCaVβ with NveCaVβ NTerm domain (plasmid) | This paper | NVE_Nterm_rat_cacnb2a | |
| Sequence-based reagent | EdCaVβ1 primer F | This paper | Custom PCR primer | GGATTTCGCCCTGAGCAA |
| Sequence-based reagent | EdCaVβ1 primer R | This paper | Custom PCR primer | TGGATCTCCGAAGGAGTTGA |
| Sequence-based reagent | EdCaVβ1 probe | This paper | Custom probe | FAM-ACATAGAACTTGATAGTCTCGAGCACG-BHQ-1 |
| Sequence-based reagent | EdCaVβ2 primer F | This paper | Custom PCR primer | TGTCCAGAGCTTCACAAAGG |
| Sequence-based reagent | EdCaVβ2 primer R | This paper | Custom PCR primer | TCTTCGTCAACACTACTTGTATCA |
| Sequence-based reagent | EdCaVβ2 probe | This paper | Custom probe | HEX-ACTCAGAACCTGCTTACAGAGCCT-BHQ-1 |
| Commercial assay or kit | One-Step RT-ddPCR Advanced Kit for Probes | Bio-Rad Laboratories | Cat #1864021 | |
| Commercial assay or kit | Droplet Gen Oil for Probes | Bio-Rad Laboratories | Cat #1863005 | |
| Chemical compound, drug | CdCl2 | Sigma | Cat #202908 | |
| Software, algorithm | Rcorrector | Song and Florea, 2015 | https://github.com/mourisl/Rcorrector | |
| Software, algorithm | FilterUncorrectablePEfastq.py | Harvard Informatics | https://github.com/harvardinformatics/TranscriptomeAssemblyTools | |
| Software, algorithm | Trimgalore | The Babraham Institute Bioinformatics Group | https://github.com/FelixKrueger/TrimGalore | |
| Software, algorithm | Bowtie2 | Langmead and Salzberg, 2012 | https://bowtie-bio.sourceforge.net/bowtie2/index.shtml | |
| Software, algorithm | Fastqc | The Babraham Institute Bioinformatics Group | https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ | |
| Software, algorithm | Trinity | Grabherr et al., 2011 | https://github.com/trinityrnaseq | |
| Software, algorithm | Transdecoder | Haas, BJ. https://github.com/TransDecoder/TransDecoder | https://github.com/TransDecoder/TransDecoder | |
| Software, algorithm | DIAMOND | Buchfink et al., 2015 | https://github.com/bbuchfink/diamond | |
| Software, algorithm | Kallisto | Bray et al., 2016 | https://github.com/pachterlab/kallisto | |
| Software, algorithm | Clustal Omega | Madeira et al., 2022 | https://www.ebi.ac.uk/Tools/msa/clustalo/ | |
| Software, algorithm | Geneious Prime | Geneious | RRID: SCR_010519 | https://www.geneious.com |
| Software, algorithm | MAFFT v.7 | Katoh and Standley, 2013 | https://mafft.cbrc.jp/alignment/server/ | |
| Software, algorithm | ModelFinder | Kalyaanamoorthy et al., 2017 | http://www.iqtree.org/ModelFinder/ | |
| Software, algorithm | IQ-TREE v2.0 | Minh et al., 2020 | http://www.iqtree.org/ | |
| Software, algorithm | UFBoot2 | Hoang et al., 2018 | https://bio.tools/ufboot2 | |
| Software, algorithm | QuantaSoft | Bio-Rad Laboratories | https://www.bio-rad.com/en-us/life-science/digital-pcr/qx200-droplet-digital-pcr-system/quantasoft-software-regulatory-edition | |
| Software, algorithm | pClamp 11 Software Suite | Molecular Devices | https://www.moleculardevices.com/products/axon-patch-clamp-system/acquisition-and-analysis-software/pclamp-software-suite | |
| Software, algorithm | MetaMorph Microscopy Automation and Image Analysis Software | Molecular Devices | https://www.moleculardevices.com/products/cellular-imaging-systems/acquisition-and-analysis-software/metamorph-microscopy | |
| Software, algorithm | Fiji (ImageJ) | Schindelin et al., 2012 | RRID: SCR_002285 | |
| Software, algorithm | GraphPad Prism | GraphPad Software | RRID: SCR_002798 | http://www.graphpad.com/ |
| Software, algorithm | Cell Sens | Olympus | https://www.olympus-lifescience.com/en/software/cellsens/ | |
| Commercial assay or kit | BaseScope Duplex Reagent Kit | Advanced Cell Diagnostics | Cat #323800 | |
| Sequence-based reagent | BaseScope Probe-BA-Epa- LOC110232233-tvX3- 1zz-st-C1 (EdCaVβ1) | Advanced Cell Diagnostics | Custom probe 1162801 C1 | |
| Sequence-based reagent | BaseScope Probe-BA-Epa- LOC110232233-tvX6- 1zz-st-C2 (EdCaVβ2) | Advanced Cell Diagnostics | Custom probe 1162811 C2 | |
| Sequence-based reagent | BaseScope Duplex Positive Control Probe-Human [Hs]-C1- Hs-PPIB-1zz/C2-POLR2A-1zz | Advanced Cell Diagnostics | Cat #700101 | |
| Sequence-based reagent | BaseScope Duplex Negative Control Probe-C1-DapB-1zz/C2- DapB-1zz | Advanced Cell Diagnostics | Cat #700141 |
Animals and cells
Request a detailed protocolStarlet sea anemones (Nematostella vectensis) were provided by the Marine Biological Laboratory (Woods Hole, Massachusetts). Adult animals of both sexes were used and kept on a 14 hr light/10 hr dark cycle at 26 °C in 1/3 natural sea water (NSW). Exaiptasia spp. were purchased through Carolina Biological Supply Company (Cat #162865). Adult animals of both sexes were used following being kept on either a 10 hr light/14 hr dark cycle at 26 °C in natural sea water (NSW) or a 14 hr light/10 hr dark cycle at 26 °C in natural sea water (NSW). Cassiopea spp. were purchased through Carolina Biological Supply Company (Cat #162936). Unless stated otherwise, all animals were fed freshly hatched brine shrimp (Artemia) twice a week.
Exaiptasia diaphana were bleached through chemical methods (menthol-induced). Menthol (100 mM in 100% ethanol; Sigma-Aldrich) was added to NSW at a final concentration of 0.2 mM (Matthews et al., 2016). The anemones were incubated in the menthol/NSW treatment solution for a maximum of 8 hr per day and outside of treatments anemones were incubated in NSW. For 2 weeks, anemones were treated 4 days per week and kept in the dark continuously starting from day 1 of treatment, aside from treatment changes. Animals were fed with Artemia approximately twice per week between bleaching treatments, enabling successful bleaching with minimal mortality. Their symbiotic status was assessed via fluorescence microscopy at the end of each week. For starvation experiments, animals were fed to excess for 1–2 weeks before the trial and withheld food entirely during the trial period and given water changes twice a week.
Nematostella nematocytes were isolated from tentacle tissue, which was harvested by anesthetizing animals in high magnesium solution containing (mM): 140 NaCl, 3.3 Glucose, 3.3 KCl, 3.3 HEPES, 40 MgCl2. Cells were isolated from tentacles immediately prior to electrophysiology experiments by treatment with 0.05% Trypsin at 37 °C for 15–20 min and mechanical dissociation in divalent free recording solution (mM): 140 NaCl, 3.3 Glucose, 3.3 KCl, 3.3 HEPES, pH 7.6. Dissociated cells were held on ice until use. Basitrichous isorhiza nematocytes were isolated from tentacles and identified by the presence of a capsule with high refractive index containing a barbed thread, oblong shape, and the presence of a cnidocil. Exaiptasia nematocytes were isolated from tentacle tissue immediately prior to electrophysiology experiments by incubation in a heat shock dissociation solution with (in mM): 430 NaCl, 10 KCl, 150 sucrose, 5 NaEGTA, 10 HEPES, 10 glucose, pH 7.6 at 45 °C for 15–20 min and mechanical dissociation in the same solution. Dissociated cells were held on ice until use. Nematocytes were isolated from tentacles and identified by the presence of a capsule with high refractive index, oblong shape, and the presence of one or multiple apical cilia.
HEK293T cells (ATCC, Cat# CRL-3216, RRID:CVCL_0063, authenticated and validated as negative for mycoplasma by vendor) were grown in DMEM, 10% fetal calf serum, and 1% penicillin/streptomycin at 37 °C, 5% CO2. For transfection, HEK293 cells were washed with Opti-MEM Reduced Serum Media (Gibco) and transfected using lipofectamine 2000 (Invitrogen/Life Technologies Cat #11668019) according to the manufacturer’s protocol. One µg each of M. musculus (mouse) cacna1a and rat cacna2d1 and one of a wide variety of beta subunits (Nematostella vectensis cacnb2.1 (NveCaVβ), Rattus norvegicus (rat) cacnb2a (rCaVβ2a), Exaiptasia diaphana CaVβs (EdCaVβ1, EdCaVβ2), Cyanea capillata CaVβ (CcCaVβ), Physalia physalis CaVβ (PpCaVβ), Clytia hemisphaerica CaVβ (ChCaVβ), Cassiopea xamachana CaVβ (CxCaVβ2)) were coexpressed with 0.5 µg eGFP. We also assayed an array of different EdCaVβ2 mutants with N-termini from different animals by coexpressing 0.5 µg eGFP, 1 µg of M. musculus (mouse) cacna1a and rat cacna2d1, and one of a variety of beta subunits (Nematostella vectensis cacnb2.1 mutant (NveCaVβ-N), R. norvegicus (rat) cacnb2a mutant (rCaVβ-N), Exaiptasia diaphana CaVβ mutants (EdCaVβ1-N, EdCaVβ2-N), Cyanea capillata CaVβ mutant (CcCaVβ-N), Physalia physalis CaVβ mutant (PpCaVβ-N), Clytia hemisphaerica CaVβ mutant (ChCaVβ-N), Cassiopea xamachana CaVβ mutant (CxCaVβ2-N)). To enhance channel expression, cells were incubated with transfection mix containing plasmid DNA and Lipofectamine 2000 in Opti-MEM for 6 hr at 37 °C. Cell were then re-plated on coverslips, incubated for 1–2 hr at 37 °C, and then incubated at 30 °C for 2–6 days before experiments. Rat cacna2d1 (RRID: Addgene_26575) and cacna1a were gifts from D. Lipscombe (RRID: Addgene_26578) and cacnb2a was a gift from A. Dolphin (RRID: Addgene_107424).
Molecular biology
Request a detailed protocolRNA was prepared from tentacles, body, and acontia tissues of WT and bleached adult Exaiptasia using published methods (Stefanik et al., 2013). Each tissue was homogenized (Millipore Sigma Cat #Z359971) and RNA was extracted using TRIzol Reagent (Thermo Fisher Cat #15596026), then after skipping the salt precipitation steps, RNA was purified and concentrated with the RNA Clean & Concentrator-5 kit (Zymo Research). For ddPCR experiments, droplet generation (QX200 Droplet Generator BioRad Cat #1864002) and transfer of droplets to ddPCR 96-Well Plates (Bio-Rad Cat #12001925) were performed according to manufacturer’s instructions (Instruction Manual, QX200 Droplet Generator – Bio-Rad). Custom primers and probes and One-Step RT-ddPCR Advanced Kit for Probes (Bio-Rad Cat #1864021) reaction reagents and Droplet Generation Oil for Probes (Bio-Rad Cat #1863005) were sourced from Bio-Rad (see Key Resources Table for primer and probe sequences). The ddPCR plate was sealed with a Pierceable Foil Heat Seal (Bio-Rad Cat #1814040) and the PX1 PCR Plate Sealer (Bio-Rad Cat #1814000). Plates were transferred to a Bio-Rad Thermalcycler C1000 (Bio-Rad Cat #1851197). The cycling protocol was the following: 45 °C reverse transcription step for 60 min, 95 °C enzyme activation step for 10 min followed by 40 cycles of a two-step cycling protocol (denaturation step of 95 °C for 30 s and annealing/extension step of 58 °C for 1 min), 98 °C enzyme deactivation step for 10 min, and holding at 12 °C for an indefinite period before transfer to the QX200 Droplet Generator. The plates were read with the Bio-Rad QX200 Droplet Generator & Reader (Cat #1864003) and the RNA concentration per sample was processed using QuantaSoft (Bio-Rad Cat #1864011). Data were exported to Microsoft Excel and Prism (Graphpad) for further statistical analysis.
Most plasmids, including Nematostella vectensis cacnb2.1 (NveCaVβ), Exaiptasia diaphana CaVβs (EdCaVβ1, EdCaVβ2), Cyanea capillata CaVβ (CcCaVβ), Physalia physalis CaVβ (PpCaVβ), Clytia hemisphaerica CaVβ (ChCaVβ), Cassiopea xamachana CaVβ (CxCaVβ2), Nematostella vectensis cacnb2.1 mutant (NveCaVβ-N), R. norvegicus (rat) cacnb2a mutant (rCaVβ-N), Exaiptasia diaphana CaVβ mutants (EdCaVβ1-N, EdCaVβ2-N), Cyanea capillata CaVβ mutant (CcCaVβ-N), Physalia physalis CaVβ mutant (PpCaVβ-N), Clytia hemisphaerica CaVβ mutant (ChCaVβ-N), Cassiopea xamachana CaVβ mutant (CxCaVβ2-N), were synthesized by Genscript (Piscataway, NJ). Sequence alignments were carried out using Clustal Omega. Wild type and Chimeric CaVβ sequences are listed in Figure 5—source data 1.
Cnidarian beta sequences were obtained from RNA sequencing or NCBI: Nematostella vectensis cacnb2.1 (NveCaVβ) sequence (Weir et al., 2020), Exaiptasia diaphana CaVβs from RNA sequencing and confirmation from NCBI accession number KXJ28099.1 (EdCaVβ1) and NCBI accession number XP_020893045.1 (EdCaVβ2), Cyanea capillata CaVβ (CcCaVβ) from NCBI accession number AAB87751.1 (Jeziorski et al., 1998), Physalia physalis CaVβ (PpCaVβ) from NCBI accession number ABD59026 (Bouchard et al., 2006), Clytia hemisphaerica CaVβ (ChCaVβ) from the MARIMBA database (Leclère et al., 2019), Cassiopea xamachana CaVβ (CxCaVβ2) from RNA sequencing as TRINITY_DN5778_c3_g1_i5.p1.
Transcriptomics
Request a detailed protocolExaiptasia were anesthetized in 15% MgCl2 NSW solution in a dish surrounded by an ice bath for 15 min. Tentacle, body, and acontia tissue were dissected and flash frozen in the presence of liquid nitrogen. Cassiopea xamachana were anesthetized in 10% MgCl2 NSW solution in a dish surrounded by an ice bath for 15–20 min. Oral arms, bell, and cassiosome tissues were dissected and flash frozen in the presence of liquid nitrogen. All tissues were stored at –80 °C until RNA extraction, library preparation, and RNA sequencing was performed by Genewiz (Azenta) using a HiSeq (2x150 bp) platform. Reads were examined for base quality distribution, kmer frequencies and adapter contamination by position in the read using fastqc (The Babraham Institute Bioinformatics Group), then where relevant, Rcorrector was used to remove erroneous k-mers (Song and Florea, 2015) and the FilterUncorrectablePEfastq python script from the Harvard Informatics group was used to discard read pairs. TrimGalore (The Babraham Institute) was then used to remove adapter contamination in reads and where relevant, Bowtie2 (Langmead and Salzberg, 2012) was used to remove reads originating from rRNA and Trinity was used to assemble reference transcriptomes de novo (Grabherr et al., 2011). Transdecoder was used to identify open reading frames (Haas, BJ) and Diamond used to annotate the transcriptome (Buchfink et al., 2015). Reads were pseudo-aligned and transcript abundance (TPM) was quantified using Kallisto (Bray et al., 2016) and our novel transcriptome assemblies as a reference and visualization and alignments were performed with Geneious Prime software and/or Clustal Omega (Madeira et al., 2022).
Electrophysiology
Request a detailed protocolRecordings were carried out at room temperature using a MultiClamp 700B amplifier (Axon Instruments) and digitized using a Digidata 1550B (Axon Instruments) interface and pClamp software (Axon Instruments). Whole-cell recording data were filtered at 1 kHz and sampled at 10 kHz. CaV activation data were leak-subtracted online using a p/4 protocol, and all membrane potentials were corrected for liquid junction potentials.
For whole-cell nematocyte recordings, borosilicate glass pipettes were polished to 8–10 MΩ for Nematostella and 4–6 MΩ for Exaiptasia, respectively. The standard Nematostella medium was used as the extracellular solution and contained (in mM): 140 NaCl, 3.3 glucose, 3.3 KCl, 3.3 HEPES, 2 CaCl2, 0.5 MgCl2, pH 7.6, 260-280mOsm. The standard Exaiptasia medium was used as extracellular solution and contained (in mM): 430 NaCl, 10 KCl, 10 CaCl2, 50 MgCl2, 10 HEPES, pH 7.6, 800-900mOsm. The intracellular solution for both Nematostella and Exaiptasia contained (in mM): isolating inward currents (in mM): 500 cesium methanesulfonate, 4 MgCl2, 10 CsEGTA, 10 HEPES, 30 sucrose, pH 7.6, 260-280mOsm for Nematostella and 800-900mOsm for Exaiptasia. For Nematostella nematocyte recordings, voltage-dependent inactivation was measured during a 200ms activating pulse of –20 mV following a series of 1 s pre-pulses ranging from –110mV to 30 mV, holding at –110 mV. Voltage-gated currents were measured through a series of 200ms voltage pulses in 10 mV increments from –110mV to 70 mV, holding at –110 mV. For Exaiptasia nematocyte recordings, voltage-dependent inactivation was measured during a 200ms activating pulse of 0 mV following a series of 1 s pre-pulses ranging from –110mV to 30 mV, holding at –110 mV. For both Nematostella and Exaiptasia, voltage-gated currents were measured through a series of 200ms steps 200ms voltage pulses in 10 mV increments from –110mV to 70 mV, holding at –110 mV. For Cd2+ experiments, 500 µM Cd2+ (dissolved in water) was applied locally and voltage-dependent activation was assessed through a single 200ms step to 0 mV from a holding potential of –110 mV.
For whole-cell recordings in HEK293 cells, pipettes were 3–6 MΩ. The standard extracellular solution contained (in mM): 140 NaCl, 5 KCl, 10 HEPES, 2 CaCl2, 2 MgCl2, 10 Glucose, pH 7.4, 300-310mOsm. The intracellular solution contained (in mM): 5 NaCl, 140 cesium methanesulfonate, 1 MgCl2, 10 EGTA, 10 HEPES, 10 sucrose, pH 7.2, 300-310mOsm. For Ca2+ currents in heterologously expressed channels, voltage-dependent inactivation was measured in one of two ways: (1) during an activating pulse of 0 mV following a series of 1 s pre-pulses ranging from –110mV to 50 mV and holding potential of –80 mV; or (2) during an activating pulse of 0 mV following a series of 1 s pre-pulses ranging from –110mV to 80 mV and holding potential of –90 mV. Voltage-gated Ca2+ currents were measured in response to 200ms voltage pulses in 10 mV increments from –130mV to 80 mV with –110 mV holding potential. Voltage-dependent inactivation was quantified as I/Imax, with Imax occurring at the voltage pulse following a –110 mV prepulse. In some instances, inactivation curves could not be fitted with a Boltzmann equation and were instead fitted with an exponential. G-V relationships were derived from I-V curves by calculating G: G=ICaV/(Vm-Erev) and fit with a Boltzmann equation. Data was processed and analyzed in Clampfit (pClamp 11 Software Suite, Molecular Devices) and Microsoft Excel and Prism (GraphPad).
In situ hybridization (BaseScope)
Request a detailed protocolAdult Exaiptasia were paralyzed in anesthetic solution (15% MgCl2), rinsed in PBS, then embedded in Tissue-Tek O.C.T. Compound (Sakura Cat #4583) in cryomolds (Sakura Tissue-Tek Cryomold, Intermediate, Cat #4566) and flash frozen on dry ice and stored at –80 °C. Cryostat sections (18–20 µm) were adhered to Fisherbrand Superfrost Plus Microscope Slides (Fisher Scientific Cat #12-550-15) and flash frozen on dry ice and stored at –80 °C until used for BaseScope. The BaseScope Duplex Detection Reagent Kit (Advanced Cell Diagnostics Cat #323800) and the manufacturer’s manual (BaseScope Duplex Detection Reagent User Manual, ACDBio) was followed to hybridize custom probes or positive control (Cat #700101) or negative control probes (Cat #700141) to targets in tissue cryosections and amplify signals. Samples were imaged on an Olympus BX41 Phase Contrast & Darkfield Microscope (Olympus Cat #BX41-PH-B) and images were acquired using the Olympus CellSens software and Olympus DP25 5MP Color Firewire Camera.
Behavior
Request a detailed protocolDischarge of nematocysts was assessed based on well-established assays (Gitter et al., 1994; Watson and Hessinger, 1994; Weir et al., 2020). For assaying discharge, 5 mm round coverslips were coated with a solution of 25% gelatin (w/v) dissolved in NSW (for Exaiptasia) or 1/3 NSW (for Nematostella) and allowed to cure 3–4 hr prior to use. Coverslips were presented to the animal’s tentacles for 5 s and then immediately imaged at ×20 magnification using a transmitted light source. To assay behavioral responses to prey-derived chemicals, freshly hatched brine shrimp were flash frozen and ground to a powder with a mortar and pestle (Fisherbrand), then filtered through a 0.22 µm syringe filter (VWR Cat #28145–501) and osmolarity adjusted for the specific anemone species. Coverslips were submerged in prey extract for 10 seconds then immediately presented to the animal. Nematocytes visualized on coverslips were only those that embedded in the gelatin after discharge. For experiments using pharmacological agents such as CdCl2, coverslips were submerged in a solution of 1 M (Exaiptasia) or 10 mM (Nematostella) Cd2+ in milli-Q water for 10 s then immediately presented to the animal. After performing the experiments, the animals were given several water changes to remove Cd2+. Experiments carried out in the absence of extracellular Ca2+ were nominally Ca2+ free and did not include use of extracellular chelators. The region of the highest density of discharged nematocytes on the coverslip was imaged at 20 X. Images were acquired with MetaMorph Microscopy Automation and Image Analysis Software (Molecular Devices) and the number of discharged nematocysts was counted by eye. Images were processed in Fiji (ImageJ) (Schindelin et al., 2012). Exaiptasia and Nematostella tentacles were examined by cutting a small portion of exposed tentacles and sandwiched between glass coverslips and then imaged at 20 X with the MetaMorph software.
Phylogenetic and genomic analyses
Request a detailed protocolTo infer exon boundaries and isoforms, we aligned EdCaVβ1 (NCBI accession number LJWW01000015.1) and EdCaVβ2 (NCBI accession number XM_021037386.2) to the Exaiptasia diaphana reference genome (BioProject PRJNA261862) (Baumgarten et al., 2015) using GMAP version 2015-07-23 (Wu and Watanabe, 2005). For phylogenetic analyses, we aligned nucleotide sequences with MAFFT v.7 (Katoh and Standley, 2013). We used ModelFinder (Kalyaanamoorthy et al., 2017) to assess the best model of substitution for phylogenetic inference. We estimated a maximum likelihood gene tree in IQ-TREE v2.0 (Minh et al., 2020). Support for clades was calculated using ultrafast bootstrap approximation UFBoot2 (Hoang et al., 2018). Percentage of identity for amino acids was calculated in overlapping windows.
Mathematical model: optimal control theory for the stinging response
Predatory stinging
Request a detailed protocolTo model Nematostella, we assume the agent stings for predation. We thus introduce the state of starvation, , that ranges from 0 to 1; at the agent is least starved and at the agent is most starved (Figure 2—figure supplement 1A top). At each time step, the agent decides to perform an action (sting), , representing the intensity of the attack; ranges from 0 to 1, and is experimentally compared to the fraction of nematocytes fired in the behavioral assay. Each action has a cost that is proportional to the fraction of nematocysts that are fired, where is the cost of discharging all nematocysts at once, or cost of full discharge, and we first consider constant. Each stinging event has a probability of achieving successful predation, where increases with (more intense attacks are more costly and more likely to succeed). A successful attack leads to the transition to the next state where the agent is more satiated whereas a failed attack leads to higher starvation state . The most starved state is absorbing, which is equivalent to a point of no return. A reward is assigned to the state of starvation reached upon attack, indicating its desirability ( is a decreasing function of ), and the most starved state entails a starvation penalty . Without loss of generality, all costs and rewards are normalized to the penalty of starvation, hence penalty of starvation is . Our goal is to choose actions that maximize the expected sum of all future net rewards (reward - cost) for each state, which is called the value function. As customary in infinite horizon problems, we ensure convergence of the value function by introducing an effective horizon, i.e. by discounting exponentially rewards that are further in the future with a discount rate .
The optimal value of a state, and the corresponding optimal action are obtained by solving the Bellman Optimality equation (Bellman, 2003), with the boundary condition
(1)
(2)
Predatory stinging increases with starvation
Request a detailed protocolWe solve Equations (1) and (2) numerically with the value iteration algorithm (Sutton, 2018) and analytically under the assumption that varies slowly with (see Asymptotics for predatory stinging). The asymptotic solution reproduces well the numerical results (compare lines and full circles in Figure 2B right, where we used and and and showcased two different functional forms for r(s), ; ). We also explored how well the asymptotic result can capture the trend of the numerical result by varying the parameters in these three different forms for the reward (Figure 2—figure supplement 2). The asymptotic solution shows that the stinging response increases with starvation under broad conditions and not only for specific forms of rewards , transitions and costs , (i.e. as long as and are concave functions and is convex, see Asymptotics for predatory stinging). To exemplify the importance of acting optimally to save resources, we considered two agents, one acting optimally and one acting randomly i.e. shooting with a number of nematocysts uniformly distributed between and . Both agents start at the same starvation state ( in Figure 2D) and use on average the same number of nematocysts, but the random agent reaches starvation typically in tens of steps, whereas the optimal agent converges to a steady state (around in the figure) and hardly ever reaches severe starvation. Predatory stinging increases with starvation even when costs increase moderately with starvation; it will eventually decrease with starvation when cost increase is exceedingly steep (see Asymptotics for predatory stinging – changing cost).
Asymptotics for predatory stinging
Request a detailed protocolShort-hand notation: . When we obtain it by zeroing the derivative with respect to in Equation 1:
(3)
Plugging Equation 3 into the Bellman Equation 1 we obtain:
(4)
From Equation 3 there is also
(5)
Equations 4 and 5 are two equations in the four unknowns and . These equations can be solved iteratively by coupling all states and using the boundary conditions on the absorbing state. However, the exact iterative solution is not particularly instructive. Instead, we will make a simplifying assumption that leads to a good approximation that can be used to gather a qualitative understanding of the prediction. Assume that varies slowly with so that (better approximations may be achieved by assuming a first order expansion). Then we obtain a third equation by writing Equation 4 for the state
(6)
We can then repeat the trick to obtain a fourth equation. To this end, we first eliminate by combining Equations 5 and 6:
(7)
(8)
Repeating the trick, we can write Equation 7 for and using that we obtain a fourth equation to close the system:
(9)
The system is now closed with the 4 Equations 4–6; 9 in the 4 unknowns and . We solve for by eliminating from Equations 4; 9 and plugging the expression for from Equation 7. After some (tedious) algebra, we obtain that satisfies the simple relation:
(10)
Both the asymptotic solution from Equation 10 and the numerical solution from value iteration are used in the main text (Figure 2B right, symbols and lines respectively). We showcase the robust match between the asymptotic and numerical solutions to Equation 1 by using a variety of functional forms of the reward function and varying the parameters (Figure 2—figure supplement 2). The asymptotics break down if abrupt changes in the rewards and transition rates are assumed, which leads to exceeding slopes in the optimal policy (data not shown). Equation 12 has a non-trivial solution when , is a decreasing function, and is an increasing function. If we additionally assume that is concave, and is a decreasing function of (for example, is strictly concave and is convex), then Equation 12 prescribes that increases with , as seen graphically in Figure 2—figure supplement 1A bottom. Hence independently of the specific functional forms of and , as long as these broad assumptions are valid, optimal stinging for predation entails more intense attacks as starvation increases. For different assumptions of reward function , cost function , and probability , we can easily substitute the specific expressions into Equation 10 and solve for for every .
Asymptotics for predatory stinging – changing cost
Request a detailed protocolWe applied a cost to predatory stinging that increases with starvation. We found numerically that predatory stinging still increases for moderate increase of with . The result is exemplified in Figure 2C using the same functional form for of the defensive stinging (Figure 2B, open circles). For a more intense increase of with predatory stinging eventually decreases with starvation (see Figure 2—figure supplement 4, for a comparison with four different cost functions from numerical solutions of Equations 1; 2).
These results can be easily understood from our asymptotic solution (10), which appears to still hold when (data not shown -- a formal proof of the asymptotic solution for this case and further consequences for Markov Decision Processes are beyond the scope of the current paper). Indeed, if increases slightly with , the light blue curve in Figure 2—figure supplement 3 slightly shifts downward with . If the shift is sufficiently small, its intersection with the green curves still occurs for increasing values of (dashed line in Figure 2—figure supplement 3). However, a dramatic increase of with will shift the light-blue curve downward considerably, and the intersection will eventually move backward (dotted line in Figure 2—figure supplement 3). In plain words, when the cost of nematocyst discharge for starved animals is dramatically larger than for well-fed animals, the benefits of predation are eventually outweighed by its cost and the animals will sting less with starvation (exemplified in Figure 2—figure supplement 4, green and yellow curves). Note that the most extreme increase of cost with starvation is unrealistic as it entails that the cost of stinging is nearly irrelevant when well fed and outweighs the benefits of feeding when starving (see green and yellow cost functions in Figure 2—figure supplement 4A). This scenario may become more relevant upon severe starvation, which we do not explore experimentally.
Defensive stinging
Request a detailed protocolTo model Exaiptasia, we assume the agent stings for defense, thus the associated Markov process models transitions between the states of safety, which we indicate with L, and danger, which we indicate with D (Figure 2—figure supplement 1A top). The state of starvation is not affected by stinging and instead is dictated by a separate process that relies on symbionts and which we do not model. Similar to the previous model, the agent chooses an action, , representing the intensity of the attack. Each attack has a likelihood to succeed and an associated cost where is the cost of full discharge of all nematocysts at once. A successful attack allows the animal to remain in state and receive a unit reward; a failed attack leads to state and penalty -1. is an absorbing state hence . The optimal value and action in state follow:
(11)
Analytic solution for defensive stinging
Request a detailed protocolZeroing the derivative of the argument on the r.h.s. of Equation 11 leads to
Here, is the probability of success of action and it ranges from to . Combining these equations we obtain an implicit algebraic equation for :
(12)
Assuming the specific form in Equation 12 leads to the constant optimal action: , where and . If is constant, there is a non-trivial optimal action as long as and clearly the optimal action does not depend on starvation (constant solution in Figure 2E right, with ; and ).
Defensive stinging – increasing cost
Request a detailed protocolStinging predators does not improve nutritional state, thus transitions among different starvation states are not modelled for defensive stinging, but instead rely heavily on symbionts. However, to capture subtle effects of starvation on defensive stinging, we note that the cost of discharging nematocysts may still depend parametrically on whatever state of starvation the agent happens to be in. In this case, and under the assumption that the cost increases with starvation, we find that optimal defensive stinging always decreases with starvation, for any functional form of . Indeed, Equation 12 with simply reads:
(13)
where is now a partial derivative with respect to . For and , the solution is where ; , and it exists for . Outside these boundaries, the solution is either ∗=0 or ∗=1 and it is obtained by comparing for these two choices of action and choosing the one that maximizes . This solution is decreasing as can be easily demonstrated by deriving with respect to . We used this solution for to match the experimental data and obtained a fit for the cost function shown in Figure 2A (empty circles).
While we discussed a specific case for the choices of and above, optimal defensive stinging decreases with in much more general conditions. Indeed, decreases or remains constant with using the same broad classes of functions discussed for predatory stinging ( is concave and is convex in ; either or can be linear in , but not both) and assuming additionally that i.e. that the cost does not decrease with starvation. Rearranging Equation 13 we note that is defined by the point where:
(14)
The l.h.s. of Equation 14 is an increasing function of as seen by deriving with respect to and using the assumptions: ; ; and . Because increases with , the intersection of the l.h.s. with the constant value occurs at lower and lower values of as increases (see graphical representation Figure 2—figure supplement 1B bottom). Thus under the same broad assumptions for the functional forms of and , stinging for predation increases with starvation, whereas stinging for defense remains constant or decreases with starvation.
Statistical analysis
Request a detailed protocolData were analyzed with Clampfit (Axon Instruments), Prism (GraphPad), or QuantaSoft (BioRad Laboratories) and are represented as mean ± sem. n represents independent experiments for the number of cells/patches or behavioral trials. Data were considered significant if P<0.05 using paired or unpaired two-tailed Student’s t-tests or one- or two-way ANOVAs. All significance tests were justified considering the experimental design and we assumed normal distribution and variance, as is common for similar experiments. Sample sizes were chosen based on the number of independent experiments required for statistical significance and technical feasibility.
Data availability
Deep sequencing data are available via the Sequence Read Archive (SRA) repository under the BioProject accession code PRJNA945904. All plasmids are available upon request. Further requests for resources and reagents should be directed to and will be fulfilled by the corresponding author, NWB (nbellono@harvard.edu). The Matlab code to obtain the optimal predicted stinging according to our Markov Decision Process is available from Zenodo.
-
NCBI BioProjectID PRJNA945904. Molecular tuning of sea anemone stinging RNA-seq.
References
-
Old cell, new trick? Cnidocytes as a model for the evolution of noveltyIntegrative and Comparative Biology 54:714–722.https://doi.org/10.1093/icb/icu027
-
The nematocyst: a molecular map of the cnidarian stinging organelleThe International Journal of Developmental Biology 56:577–582.https://doi.org/10.1387/ijdb.113472ab
-
SoftwareDynamic programmingDover.
-
Cloning and functional expression of voltage-gated ion channel subunits from cnidocytes of the Portuguese Man O’War Physalia physalisThe Journal of Experimental Biology 209:2979–2989.https://doi.org/10.1242/jeb.02314
-
Near-optimal probabilistic RNA-seq quantificationNature Biotechnology 34:525–527.https://doi.org/10.1038/nbt.3519
-
Fast and sensitive protein alignment using DIAMONDNature Methods 12:59–60.https://doi.org/10.1038/nmeth.3176
-
The ß subunit of voltage-gated Ca2+ channelsPhysiological Reviews 90:1461–1506.https://doi.org/10.1152/physrev.00057.2009
-
The calcium channel beta2 (CACNB2) subunit repertoire in teleostsBMC Molecular Biology 9:38.https://doi.org/10.1186/1471-2199-9-38
-
Calcium- and voltage-dependence of nematocyst discharge in Hydra vulgarisJournal of Comparative Physiology A 175:115–122.https://doi.org/10.1007/BF00217442
-
Full-length transcriptome assembly from RNA-Seq data without a reference genomeNature Biotechnology 29:644–652.https://doi.org/10.1038/nbt.1883
-
Alternative splicing of the beta 4 subunit has alpha1 subunit subtype-specific effects on Ca2+ channel gatingThe Journal of Neuroscience 22:1573–1582.https://doi.org/10.1523/JNEUROSCI.22-05-01573.2002
-
UFBoot2: improving the ultrafast bootstrap approximationMolecular Biology and Evolution 35:518–522.https://doi.org/10.1093/molbev/msx281
-
An option space for early neural evolutionPhilosophical Transactions of the Royal Society of London. Series B, Biological Sciences 370:20150181.https://doi.org/10.1098/rstb.2015.0181
-
Cloning and functional expression of a voltage-gated calcium channel alpha1 subunit from jellyfishThe Journal of Biological Chemistry 273:22792–22799.https://doi.org/10.1074/jbc.273.35.22792
-
The behavioral and developmental physiology of nematocystsCanadian Journal of Zoology 80:1772–1794.https://doi.org/10.1139/z02-135
-
MAFFT multiple sequence alignment software version 7: improvements in performance and usabilityMolecular Biology and Evolution 30:772–780.https://doi.org/10.1093/molbev/mst010
-
Fast gapped-read alignment with Bowtie 2Nature Methods 9:357–359.https://doi.org/10.1038/nmeth.1923
-
The genome of the jellyfish Clytia hemisphaerica and the evolution of the cnidarian life-cycleNature Ecology & Evolution 3:801–810.https://doi.org/10.1038/s41559-019-0833-2
-
Alternative splicing in the voltage-sensing region of N-Type CaV2.2 channels modulates channel kineticsJournal of Neurophysiology 92:2820–2830.https://doi.org/10.1152/jn.00048.2004
-
Search and sequence analysis tools services from EMBL-EBI in 2022Nucleic Acids Research 50:W276–W279.https://doi.org/10.1093/nar/gkac240
-
Menthol-induced bleaching rapidly and effectively provides experimental aposymbiotic sea anemones (Aiptasia sp.) for symbiosis investigationsThe Journal of Experimental Biology 219:306–310.https://doi.org/10.1242/jeb.128934
-
Corrigendum to: IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic eraMolecular Biology and Evolution 37:1530–1534.https://doi.org/10.1093/molbev/msaa131
-
Estimating the daily contribution of carbon from zooxanthellae to coral animal respiration1Limnology and Oceanography 26:601–611.https://doi.org/10.4319/lo.1981.26.4.0601
-
Nanosecond-scale kinetics of nematocyst dischargeCurrent Biology 16:R316–R318.https://doi.org/10.1016/j.cub.2006.03.089
-
Novel CaV2.1 clone replicates many properties of Purkinje cell CaV2.1 currentThe European Journal of Neuroscience 26:2950–2961.https://doi.org/10.1111/j.1460-9568.2007.05912.x
-
Fiji: an open-source platform for biological-image analysisNature Methods 9:676–682.https://doi.org/10.1038/nmeth.2019
-
The Rise and Fall of TRP-N, an Ancient Family of Mechanogated Ion Channels, in MetazoaGenome Biology and Evolution 7:1713–1727.https://doi.org/10.1093/gbe/evv091
-
A genomic view of 500 million years of cnidarian evolutionTrends in Genetics 27:7–13.https://doi.org/10.1016/j.tig.2010.10.002
-
The bioenergetics of symbiotic sea anemones (Anthozoa: Actiniaria)Symbiosis 5:103–142.
-
Control of cnida discharge: I. evidence for two classes of chemoreceptorThe Biological Bulletin 174:163–171.https://doi.org/10.2307/1541783
-
Evidence for calcium channels involved in regulating nematocyst dischargeComparative Biochemistry and Physiology Part A 107:473–481.https://doi.org/10.1016/0300-9629(94)90028-0
-
The cell biology of nematocystsInternational Review of Cytology 156:275–300.https://doi.org/10.1016/s0074-7696(08)62256-1
-
Poly(gamma-glutamic acid)s are the major constituents of nematocysts in Hydra (Hydrozoa, Cnidaria)The Journal of Biological Chemistry 265:9664–9669.
-
Neural pathways and innervation of cnidocytes in tentacles of sea anemonesHydrobiologia 530–531:117–121.https://doi.org/10.1007/s10750-004-2678-0
Article and author information
Author details
Nicholas W Bellono

Funding
National Institutes of Health (R35GM142697)
- Nicholas W Bellono
European Research Council (101002724 RIDING)
- Agnese Seminara
Air Force Office of Scientific Research (FA8655-20-1-7028)
- Agnese Seminara
National Institutes of Health (R01DC018789)
- Agnese Seminara
New York Stem Cell Foundation
- Nicholas W Bellono
Searle Scholars Program
- Nicholas W Bellono
National Science Foundation (Graduate Research Fellowship Program)
- Lily S He
National Science Foundation (Physics of Living Systems (PoLS) Graduate Fellowship)
- Lily S He
Harvard University (Simmons Award at the Harvard Center for Biological Imaging)
- Lily S He
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank B Walsh and P Kilian for assistance with animal husbandry, A Whipple and D Loftus with guidance for ddPCR experiments, A Grearson for illustrations and photographs, K Koenig and M Martindale, and the Marine Biological Laboratory for providing animals. We also thank the Harvard Center for Biological Imaging (RRID:SCR_018673), Histology Core at the Harvard Department of Stem Cell and Regenerative Biology, and The Bauer Core Facility at Harvard University for infrastructure and experimental support. This research was supported by grants to NWB from the New York Stem Cell Foundation, Searle Scholars Program, and the NIH (R35GM142697), fellowships to LH from NSF Graduate Research Fellowship Program and Physics of Living Systems (PoLS) Graduate Fellowship and the Simmons Award at the Harvard Center for Biological Imaging, grants to AS from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No 101002724 RIDING), the Air Force Office of Scientific Research under award number FA8655-20-1-7028, and the National Institutes of Health (NIH) under award number R01DC018789.
Version history
- Sent for peer review:
- Preprint posted:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.88900. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2023, He et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,971
- views
-
- 217
- downloads
-
- 6
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 2
- citations for umbrella DOI https://doi.org/10.7554/eLife.88900
-
- 4
- citations for Version of Record https://doi.org/10.7554/eLife.88900.3
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Molecular tuning of sea anemone stinging
eLife 12:RP88900.
https://doi.org/10.7554/eLife.88900.3
Further reading
-
Some sea anemones sting under certain conditions, whereas others sting indiscriminately.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}