Action does not enhance but attenuates predicted touch
- Department of Neuroscience, Karolinska Institute, Sweden
- Donders Institute for Brain, Cognition and Behaviour, Radboud University, Netherlands
Abstract
Dominant motor control theories propose that the brain predicts and attenuates the somatosensory consequences of actions, referred to as somatosensory attenuation. Support comes from psychophysical and neuroimaging studies showing that touch applied on a passive hand elicits attenuated perceptual and neural responses if it is actively generated by one’s other hand, compared to an identical touch from an external origin. However, recent experimental findings have challenged this view by providing psychophysical evidence that the perceived intensity of touch on the passive hand is enhanced if the active hand does not receive touch simultaneously with the passive hand (somatosensory enhancement) and by further attributing attenuation to the double tactile stimulation of the hands upon contact. Here, we directly contrasted the hypotheses of the attenuation and enhancement models regarding how action influences somatosensory perception by manipulating whether the active hand contacts the passive hand. We further assessed somatosensory perception in the absence of any predictive cues in a condition that turned out to be essential for interpreting the experimental findings. In three pre-registered experiments, we demonstrate that action does not enhance the predicted touch (Experiment 1), that the previously reported ‘enhancement’ effects are driven by the reference condition used (Experiment 2), and that self-generated touch is robustly attenuated regardless of whether the two hands make contact (Experiment 3). Our results provide conclusive evidence that action does not enhance but attenuates predicted touch and prompt a reappraisal of recent experimental findings upon which theoretical frameworks proposing a perceptual enhancement by action prediction are based.
Editor's evaluation
While decades of research findings have supported the idea that action attenuates predicted touch, a recent finding has countered this, proposing that action actually enhances predicted touch and that the previously observed attenuation is due to tactile contact. The present study rigorously probes these alternative hypotheses with three pre-registered experiments. They show that previous enhancement findings are due to the lack of a proper baseline condition of externally generated sensation with which to compare. In light of the recent opposing findings, the current paper is important and presents compelling evidence in favor of the sensory attenuation theory.
https://doi.org/10.7554/eLife.90912.sa0Introduction
Dominant motor control theories propose that the brain uses an internal forward model in combination with a copy of the motor command (efference copy) to predict the sensory consequences of our movements (McNamee and Wolpert, 2019; Shadmehr et al., 2008; Wolpert and Flanagan, 2001). For example, the brain predicts the upcoming touch as one reaches toward an object. These predictions allow for the correction of motor errors without relying on sensory feedback that suffers from intrinsic delays (Shadmehr et al., 2010), thereby improving the estimation of the current state of our body by combining the predicted touch with the actual sensory input (Scott, 2004; Shadmehr et al., 2008; Todorov and Jordan, 2002). These predictions attenuate the perception of the self-generated input (sensory reafference) compared to that of externally generated input (Davidson and Wolpert, 2005; Franklin and Wolpert, 2011; Wolpert and Kawato, 1998) and allow to infer whether the cause of the sensory input is the self or the environment (Brown et al., 2013; Corlett, 2020; Idei et al., 2022). A classic example of this attenuation is that we are unable to tickle ourselves with our own touch, yet we are easily tickled by the touch of others (Blakemore et al., 2000a). The attenuation of sensory reafference – also referred to as sensory cancelation – is thought to be necessary to compensate for the limited capacity of the sensory systems by optimally prioritizing the perception of more informative externally generated stimuli (Bays and Wolpert, 2008; McNamee and Wolpert, 2019). Thus, the attenuation model proposes that we dampen perceptual representations of expected self-generated stimuli to reduce redundancy and to highlight behaviorally relevant unexpected externally generated stimuli.
In contrast, an alternative theoretical framework proposes that predictions, including those arising from our motor commands, should not attenuate but instead enhance sensory signals, thereby allowing for sharper (i.e. more accurate) representations of predicted compared to unpredicted sensory events (Press et al., 2020; Press and Yon, 2019; Yon et al., 2021). This enhancement account – also referred to as the sharpening account – posits that predictions based on our motor commands are equivalent to expectations formed by statistical regularities in sensory input or from prior knowledge (e.g. at the North Pole, one expects to see a polar bear rather than an elephant) and that these predictions should bias our perception towards our expectations. The proposal mainly stems from experimental research outside the domain of action and argues that weak, noisy, or ambiguous sensory input that is in line with prior expectations should be enhanced to achieve, on average, more accurate representations. For example, we are more biased to report the presence of visual events that are statistically likely to occur rather than unlikely events (Chalk et al., 2010; Wyart et al., 2012), more sensitive to low-level visual features that are in line with prior expectations (Stein and Peelen, 2015; Teufel et al., 2018), and show greater biases when perceiving visual events that are congruent with our expectations (Hudson et al., 2016a; Hudson et al., 2016b). Such effects are thought to result from mechanisms that increase the ‘gain’ of expected information by altering the weights of different sensory signals (Summerfield and de Lange, 2014). Thus, the enhancement model proposes that we amplify the perceptual representations of expected compared to unexpected sensory input.
In the somatosensory domain, evidence supporting the attenuation model has repeatedly shown that touch delivered to one (passive) hand by the other (active) hand (i.e. self-generated touch) is perceived as weaker or less ticklish (Asimakidou et al., 2022; Bays et al., 2006; Bays et al., 2005; Blakemore et al., 1999; Kilteni et al., 2021; Kilteni et al., 2020; Kilteni et al., 2019; Kilteni et al., 2018; Kilteni and Ehrsson, 2022; Kilteni and Ehrsson, 2017a; Kilteni and Ehrsson, 2017b; Knoetsch and Zimmermann, 2021; McNaughton et al., 2023; Weiskrantz et al., 1971; Wolpe et al., 2016) and evokes attenuated somatosensory cortical activation (Blakemore et al., 1998a; Hesse et al., 2010; Kilteni and Ehrsson, 2020; Shergill et al., 2014; Shergill et al., 2013) compared to a touch of identical intensity applied on the passive hand that is externally generated. In contrast, evidence supporting the enhancement model has shown that the action of the active hand results in an increase in the perceived intensity of touch on the passive hand, provided that the active hand never receives touch simultaneously with the passive hand (i.e. hands do not make contact) (Thomas et al., 2022). This enhancement finding has been recently used to support the sharpening model and to argue that attenuation effects are due to unspecific gating processes caused by the simultaneous double tactile stimulation of the two hands rather than action prediction (Press et al., 2023).
The attenuation (or cancellation) and enhancement (or sharpening) models present strikingly different hypotheses regarding how action influences the perception of sensory input and are supported by contradictory experimental evidence, leading to debates between researchers (Corlett, 2020; Fuehrer et al., 2022; Kilteni and Ehrsson, 2022; Press et al., 2023; Press et al., 2020; Thomas et al., 2022). Clarifying how predictions about the sensory consequences of our movements affect our perception is fundamental to understanding the interaction between perception and action (McNamee and Wolpert, 2019; Shadmehr et al., 2008; Wolpert and Flanagan, 2001) but also for clinical and neurobiological theories of psychosis spectrum disorders, such as schizophrenia (Blakemore et al., 2000b; Corlett et al., 2019; Frith, 2005a; Frith, 2005b; Frith, 2019; Frith et al., 2000; Shergill et al., 2014; Shergill et al., 2005) and schizotypy (Asimakidou et al., 2022), as well as functional movement disorders (Pareés et al., 2014), Parkinson’s disease (Wolpe et al., 2018), and aging (Wolpe et al., 2016).
The present study aimed to contribute to this debate by directly contrasting the enhancement and attenuation models within the same paradigm. To this end, the same force discrimination task employed in earlier studies reporting attenuation (Asimakidou et al., 2022; Bays et al., 2006; Bays et al., 2005; Kilteni et al., 2023; Kilteni et al., 2021; Kilteni et al., 2020; Kilteni et al., 2019; Kilteni and Ehrsson, 2022; Timar et al., 2023) and enhancement (Thomas et al., 2022) was used to determine (a) whether movement of the right hand enhances the perceived magnitude of touch applied on the left hand when the two hands do not make contact (Experiments 1 and 2) and (b) whether attenuation effects are due to double tactile stimulation caused by the contact of the two hands or action prediction (Experiment 3). Capitalizing on the fact that any conclusion about whether action prediction ‘attenuates’ or ‘enhances’ the perception of the somatosensory input needs to be made with a comparison to one’s somatosensory perception in the absence of action, we also included a condition in which participants passively received externally generated touch, with which we compared the participants’ perception in all experimental conditions. Consequently, if participants perceive a touch as less or more intense during action than in the absence of action, we can infer that the received touch was attenuated or enhanced, respectively. This is a critical methodological novelty compared to previous studies (Bays et al., 2006; Thomas et al., 2022) because if such baseline conditions are missing, the same patterns of results can be incorrectly attributed to ‘attenuation’ or ‘enhancement’: for example, if one condition produces less attenuation than another, it may be interpreted as enhancement, and vice versa. Indeed, it was this baseline condition that turned out to be critical for distinguishing between attenuation and enhancement effects. All studies and analysis plans were pre-registered on the Open Science Framework prior to data collection (Methods).
Results
Experiment 1. Action does not enhance predicted touch
Thirty naïve participants moved their right index finger towards their left index finger to generate the touch on their left index finger with (contact condition) or without (no-contact condition) simultaneous stimulation on their active finger. A baseline condition in which the participants did not move their right index finger and received touch on the left index finger passively (externally generated touch) was included in order to distinguish between effects of attenuation or enhancement (Figure 1A).
Figure 1 with 1 supplement see all
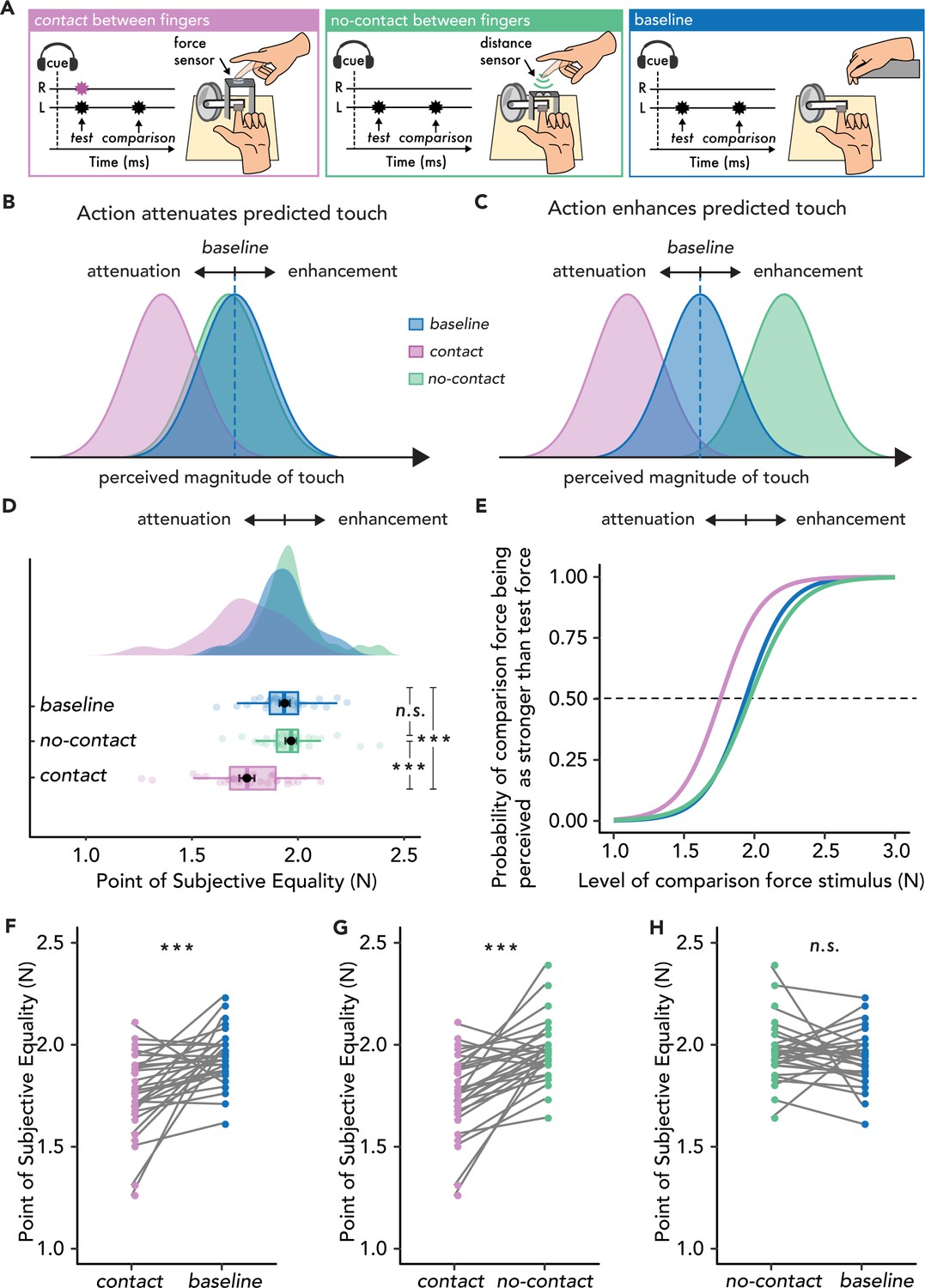
Experimental methods, hypotheses, and results of Experiment 1 (n=30).
(A) In the contact condition (magenta), participants tapped with their right index finger ‘R’ on a force sensor placed above the probe that delivered a test force of 2N to their left index finger ‘L’, followed by a comparison force randomly varying between 1 and 3N. In the no-contact condition (green), participants approached a distance sensor with their right index finger, which triggered the test force on their left index finger, thus receiving no touch on their active finger. In the baseline condition (blue), participants relaxed both hands and passively received the forces on their left index finger. (B) Hypotheses based on the attenuation model. Touch in the contact condition (magenta) should be perceived as weaker than in the baseline (blue), but touch in the no-contact condition (green) should be perceived similarly to that in the baseline (blue). (C) Hypotheses based on the enhancement model. Touch in the no-contact condition (green) should be perceived as stronger than in the baseline (i.e. enhanced) (blue), but touch in the contact condition (magenta) should be perceived as weaker than the baseline (blue). Note that attenuation effects in the contact condition are predicted both by the attenuation and the enhancement model with the difference that the attenuation model attributes these effects to action prediction, while the enhancement model attributes these effects to the simultaneous touch on the active hand. (D–H) Data are color-coded per condition. (D) Box plots show the median and interquartile ranges for the point of subjective equality (PSE) values per condition, black circles and error bars show the mean PSE ± SEM, and the raincloud plots show the individual PSE values and their distributions. No enhancement effects were observed in the no-contact condition. (E) Group psychometric functions for each condition. The leftward shift of the curve in the contact condition indicates attenuated PSE values compared to the other two conditions. (F–H). Line plots for individual participant PSE values illustrate significantly lower PSEs for the contact versus baseline (F) and no-contact (G) conditions, but no significant differences in the PSE values between no-contact and baseline (H). (***p<.001, n.s. non-significant).
During the force-discrimination task, participants judged the intensity of a ‘test’ force and a ‘comparison’ force (100 ms duration each) separated by a random interval ranging from 800 ms to 1200 ms. The forces were delivered to the pulp of the left index finger by a cylindrical probe attached to a lever driven by a DC electric motor. In each trial, participants reported which force they felt was stronger (the test or the comparison). The intensity of the test force was fixed at 2 N, while the intensity of the comparison force was systematically varied among seven force levels (1, 1.5, 1.75, 2, 2.25, 2.5, or 3 N). In the contact condition, participants moved their right index finger towards their left index finger after an auditory cue and actively tapped on a force sensor placed on top of, but not in direct contact with, the probe. The participant’s active tap on the force sensor triggered the test force on their left index finger, thereby producing simultaneous stimulation of both fingers in the contact condition and the sensation of pressing with the right index finger against the left index finger through a rigid object. Participants were told that they would always make contact between their fingers (through the probe) in this condition. In the no-contact condition, following the same auditory cue, participants moved their right index finger towards their left index finger. At the beginning of the condition, the force sensor was removed and replaced with a distance sensor that detected the relative distance of their active finger as it approached their left index finger to trigger the test force, similar to Bays et al., 2006 and Thomas et al., 2022. The distance threshold was set such that the position of the right index finger when triggering the test force was equivalent to that in the contact condition. Participants were told that they would never make contact between their fingers. Before the experiment, participants were trained to make similar movements with their right index finger in the contact and no-contact conditions. Finally, in the baseline condition (externally generated touch), participants were told to relax their right hand, and each trial began with the same auditory cue followed by the test force delivered to the participants’ left index finger 800 ms after the cue. In all trials, the comparison force was delivered after the test force, independently of the right hand, and participants verbally reported their judgment (Methods).
Participants’ responses were fitted with logistic psychophysics curves, and we extracted the point of subjective equality (PSE), which represents the intensity of the comparison force at which the test force feels equally as strong (Figure 1—figure supplement 1). Consequently, a PSE in a movement condition (contact or no-contact) that is lower than the PSE of the baseline condition indicates attenuation, while a PSE that is higher than the PSE of the baseline condition indicates enhancement (Methods). According to the attenuation model (Figure 1B), attenuation of the somatosensory input on the left hand in the contact condition compared to the baseline condition should be observed (i.e. lower PSEs) due to the test force being a predicted consequence of the action of the right hand. However, regarding the no-contact condition, earlier studies have shown that the mere movement of one hand is not sufficient to produce predictions of somatosensory input simultaneously applied on the other hand (Bays et al., 2005; Kilteni et al., 2018; Kilteni and Ehrsson, 2020; Kilteni and Ehrsson, 2017a; Kilteni and Ehrsson, 2017b; Shergill et al., 2003; Wolpe et al., 2018). Instead, a bimanual sensorimotor context is needed such as during bimanual object manipulation and bimanual contact of the hands (Blakemore et al., 1998b). In the no-contact condition, the efference copy and the sensory input are inconsistent with the sensorimotor context of touching oneself; in this case, the context of applying force on one’s finger with another finger. In other words, the attenuation model suggests that not every voluntary movement will attenuate upcoming sensory input but a naturalistic sensorimotor mapping between the movement and the sensory input is needed. Based on this, neither attenuation nor enhancement should be observed in the no-contact condition (i.e. no change in PSEs from baseline). In contrast, according to the enhancement model, if action enhances the received sensation (Thomas et al., 2022), then higher PSEs in the no-contact condition compared to the baseline condition should be observed (Figure 1C). Finally, since the enhancement model proposes that attenuation effects are attributed to unspecific (nonpredictive) gating effects caused by the simultaneous tactile stimulation of the two hands, touch in the contact condition should also be perceived as weaker, albeit not due to action prediction. In summary, Experiment 1 explicitly assessed whether action enhances the received touch when the index fingers of the two hands do not make contact.
The results showed a robust attenuation of the perceived touch when the two fingers made contact (contact condition) (Figure 1D, E and F): the PSEs were significantly lower in the contact condition than in the baseline condition (W = 422.00, p<0.001, rrb = 0.82, CI95 = [0.08, 0.25]). Similarly, the PSEs were significantly lower in the contact condition than in the no-contact condition (W = 441.00, p<0.001, rrb = 0.90, CI95 = [0.13, 0.26]) (Figure 1D, E and G). Critically, however, in the comparison that contrasts the hypotheses of the two models, the no-contact condition did not produce any significant change in the perceived magnitude of the touch compared to the baseline condition (W = 187.00, p=0.360, rrb = –0.20, CI95 = [–0.08, 0.030]) (Figure 1D, E and H). A Bayesian factor analysis provided moderate support for the absence of any difference (BF01 = 3.58). We also extracted the just noticeable difference (JND) from the psychophysical fits, which represents the participants’ force discrimination capacity between conditions. In Experiment 1, the JNDs did not significantly differ between any of the conditions (all p>0.05).
In summary, we used both frequentist and Bayesian statistics and found no evidence that the action of the right index finger produces an enhancement of the touch received on the left index finger when the fingers did not make contact, relative to the baseline. Thus, Experiment 1 does not support the hypothesis of the enhancement model that action enhances predicted touch but supports the hypothesis of the attenuation model.
Experiment 2. Previous ‘enhancement’ effects are driven by the baseline used
Having found no evidence for somatosensory enhancement in Experiment 1, Experiment 2 aimed to understand the potential source of the previously reported enhancement effects (Thomas et al., 2022). One critical methodological difference between Experiment 1 and previous evidence for enhancement concerns the baseline condition, as Thomas et al., 2022 compared the participants’ perception of the no-contact condition to a condition where the participants prepared their right index movement but received a NOGO cue to inhibit the movement, and it was that comparison that revealed enhancement effects. However, motor inhibition (i.e. planning the hand movement but not executing it) can lead to suppression of somatosensory input both on the hand that is planned to move (Hoshiyama and Sheean, 1998; Voss et al., 2008; Walsh and Haggard, 2007) and the hand that would receive touch if the movement was executed (Kilteni et al., 2018); moreover, such conditions result in a competition for attentional resources to inhibit the motor response with processes that encode sensory stimuli into memory (Chiu and Egner, 2015a; Yebra et al., 2019) (e.g. the test force in this paradigm). Therefore, if the motor inhibition condition used by Thomas et al., 2022 results in a suppression of the perceived touch, a comparison of the no-contact condition with such a baseline would produce an apparent enhancement effect.
Thirty new naïve participants participated in Experiment 2, which included a block of contact trials and a block of no-contact trials in which participants moved their right index finger, but each block also contained randomly intermixed NOGO trials where participants were cued to withhold their movement, identical to the design of Thomas et al., 2022 (Figure 2A, Figure 2—figure supplement 1). Participants were trained to make similar movements with their right index finger in the contact and no-contact trials. An externally generated touch condition was also included as an action-free baseline condition (i.e. no action execution or inhibition). If the previously reported enhancement effects are due to the baseline used, attenuated perception of touch during the motor inhibition condition (i.e. NOGO trials) should be found compared to the baseline, which would then lead to apparent enhancement effects upon comparison with the no-contact trials (Figure 2B and C) (Methods).
Figure 2 with 1 supplement see all
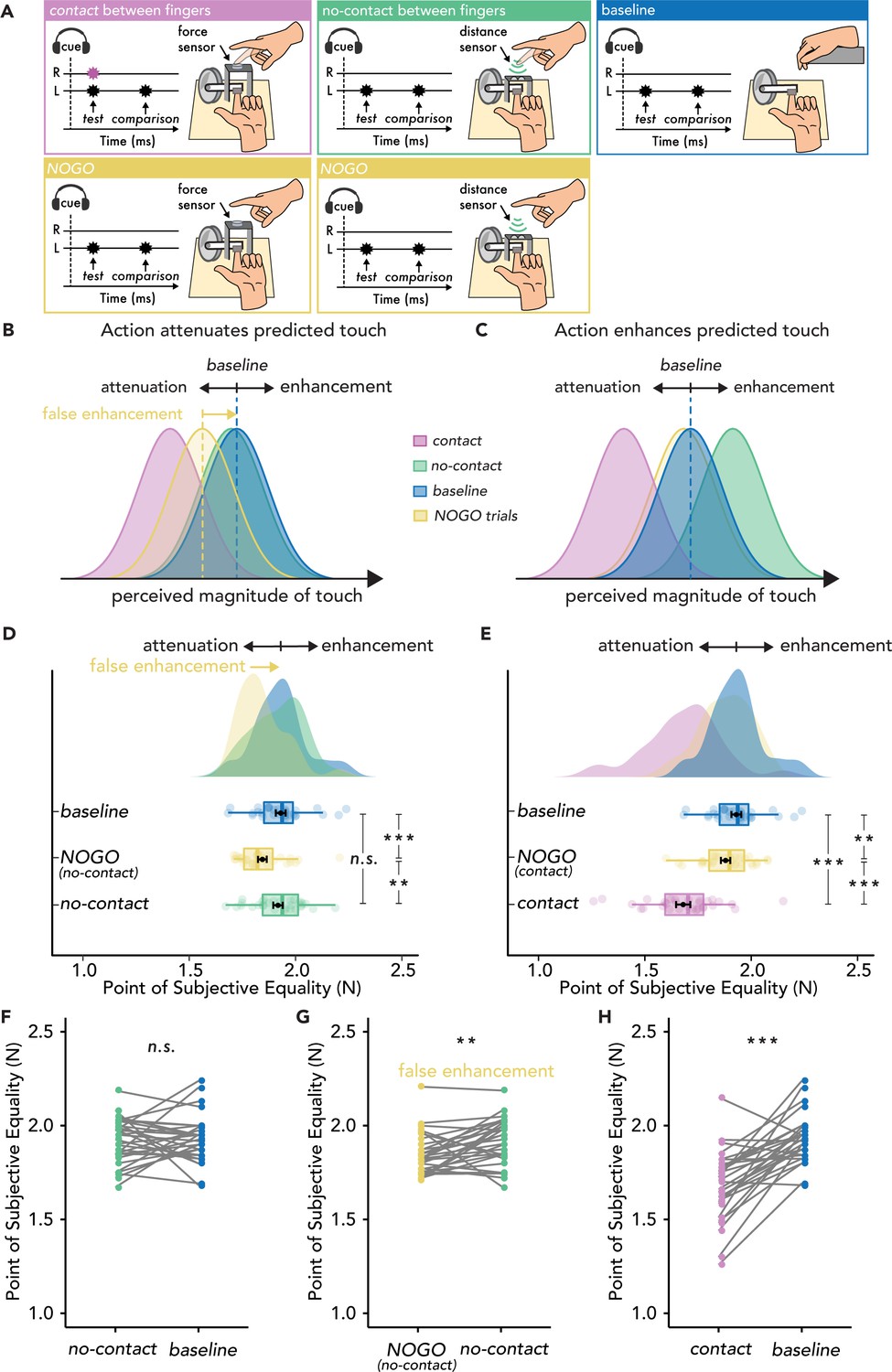
Experimental methods, hypotheses, and results of Experiment 2 (n=30).
(A) The contact (magenta) and no-contact (green) trials were identical to those of Experiment 1, with the only difference that in 50% of the trials, the participants had to inhibit their movement (NOGO trials - yellow), and the test force was delivered automatically. The baseline condition (blue) was identical to that of Experiment 1. (B) Hypotheses based on the attenuation model. Touch in the contact condition (magenta) should be perceived as weaker than in the baseline (blue), but touch in the no-contact condition (green) should be perceived similarly to that in the baseline (blue). Critically, touch may also be perceived as weaker than baseline in the NOGO trials, resulting in a ‘false enhancement’ of the no-contact trials. (C) Hypotheses based on the enhancement model. Touch in the no-contact condition (green) should be perceived as stronger than in the baseline (i.e. enhanced) (blue), but touch in the contact condition (magenta) should be perceived as weaker than the baseline (blue). (D–H) Data are color-coded per condition. (D–E) Box plots show the median and interquartile ranges for the point of subjective equality (PSE) values in the baseline and no-contact blocks (D) and in the baseline and contact blocks (E). Black circles and error bars show the mean PSE ± SEM, and the raincloud plots show the individual PSE values and their distributions. (F–H) Line plots for individual participant PSE values illustrating no significant differences in the PSE values between no-contact and baseline (F), significantly higher PSEs for the no-contact versus NOGO trials in the same block (G) and significantly lower PSEs in the contact versus baseline trials (H). (**p<0.01, ***p<0.001, n.s. non-significant).
This hypothesis (Figure 2B) was confirmed. First, all the effects of Experiment 1 were replicated in the new sample (Figure 2D, E and F & H). The contact trials yielded significant attenuation (i.e. lower PSEs) compared to the baseline condition (t(29) = 6.79, p<0.001, d = 1.24, CI95 = [0.18, 0.33]) (Figure 2E & H). Once again, there was no enhancement in the no-contact trials compared to the baseline condition (t(29) = 0.45, p=0.658, d = 0.08, CI95 = [–0.05, 0.07]) (Figure 2D & F), and the Bayesian analysis again yielded moderate evidence for the absence of any effects (BF01 = 4.69). Importantly, the PSEs in the NOGO (motor inhibition) trials were significantly lower than the baseline condition both for the contact NOGO (t(29) = 2.99, p=0.006, d = 0.55, CI95 = [0.02, 0.09]) and the no-contact NOGO trials (t(29) = 4.44, p<0.001, d = 0.81, CI95 = [0.05, 0.13]) (Figure 2D and E & G). This demonstrates that NOGO trials resulted in a suppression of perceived touch on the left hand. Critically, this led to an apparent increase in the PSE of the no-contact trials compared to the NOGO trials in the no-contact block (t(29) = –2.98, p=0.006, d = –0.54, CI95 = [–0.12,–0.02]), mimicking an ‘enhancement’ effect. Finally, PSEs were significantly lower in the contact trials than in the NOGO trials (t(29) = 5.91, p<0.001, d = 1.08, CI95 = [0.13, 0.27]), while the NOGO trials in the contact and no-contact blocks did not significantly differ (t(29) = 1.58, p=0.126, d = 0.29, CI95 = [–0.01, 0.08], BF01 = 1.697). In Experiment 2, the JNDs were not significantly different between conditions (p>0.05), except in the contact condition, where the JNDs were significantly higher than in the baseline condition (t(29) = –2.52, p=0.017, d = –0.46, CI95 = [–0.06,–0.01]).
In summary, identical to Experiment 1, we used both frequentist and Bayesian statistics and did not find any evidence that the action of the right index finger produces an enhancement of the touch received on the left index finger when the fingers do not make contact, relative to the baseline. Moreover, we showed that the purported ‘enhancement’ effect (Thomas et al., 2022) is driven by a suppression of the perceived intensity of touch following a cue to inhibit the planned movement, both relative to the baseline and to the condition where the fingers do not make contact (no-contact). Thus, Experiment 2 does not support the hypothesis that action enhances predicted touch.
Experiment 3. Action attenuates predicted touch, even without simultaneous stimulation of the active hand
Within the framework of internal forward models, the absence of attenuation in the no-contact conditions of Experiments 1 and 2 is expected, given the lack of a sensorimotor context conducive to perceiving the touch as self-generated: when contact is never made, the forces applied on the passive left hand are only arbitrarily, and not causally, associated with the movement of the active right hand. However, if there is an expectation of contact between the hands, regardless of whether the contact is eventually made, touch should be attenuated as previously shown (Bays et al., 2006). Alternatively, it could be argued that the absence of attenuation in the no-contact conditions is caused by the lack of simultaneous tactile stimulation of the active hand rather than by predictive mechanisms.
In Experiment 3, this hypothesis was explicitly tested with thirty new naïve participants (Figure 3—figure supplement 1). The same contact and no-contact conditions were included as in Experiment 1, but their relative frequency was manipulated within the same block (contact trials: 80%, no-contact trials: 20%) (Figure 3A). The force sensor the participants tapped in the contact trials was now attached to a platform that could be automatically retracted. In the contact trials, participants tapped the force sensor to trigger the test force identically to Experiments 1 and 2, but in the no-contact trials, the platform automatically retracted before trial onset, unbeknownst to the participants, leading them to unexpectedly miss the contact with the sensor but still trigger the test force only by the position of their right index finger. Participants’ vision was occluded so that they could not know whether the next trial would be a contact or a no-contact trial. According to the attenuation model, providing a bimanual sensorimotor context in 80% of the trials should lead participants to form predictions about the somatosensory consequences of their movement in most of the trials and thus attenuate the received touch on their passive left hand compared to the baseline, even if the touch of their active hand was unexpectedly missed (Figure 3B). In contrast, if attenuation is a nonpredictive process caused by the mere simultaneous tactile stimulation of the active finger, no attenuation effects in the no-contact trials should be observed with respect to the baseline (Figure 3C).
Figure 3 with 3 supplements see all
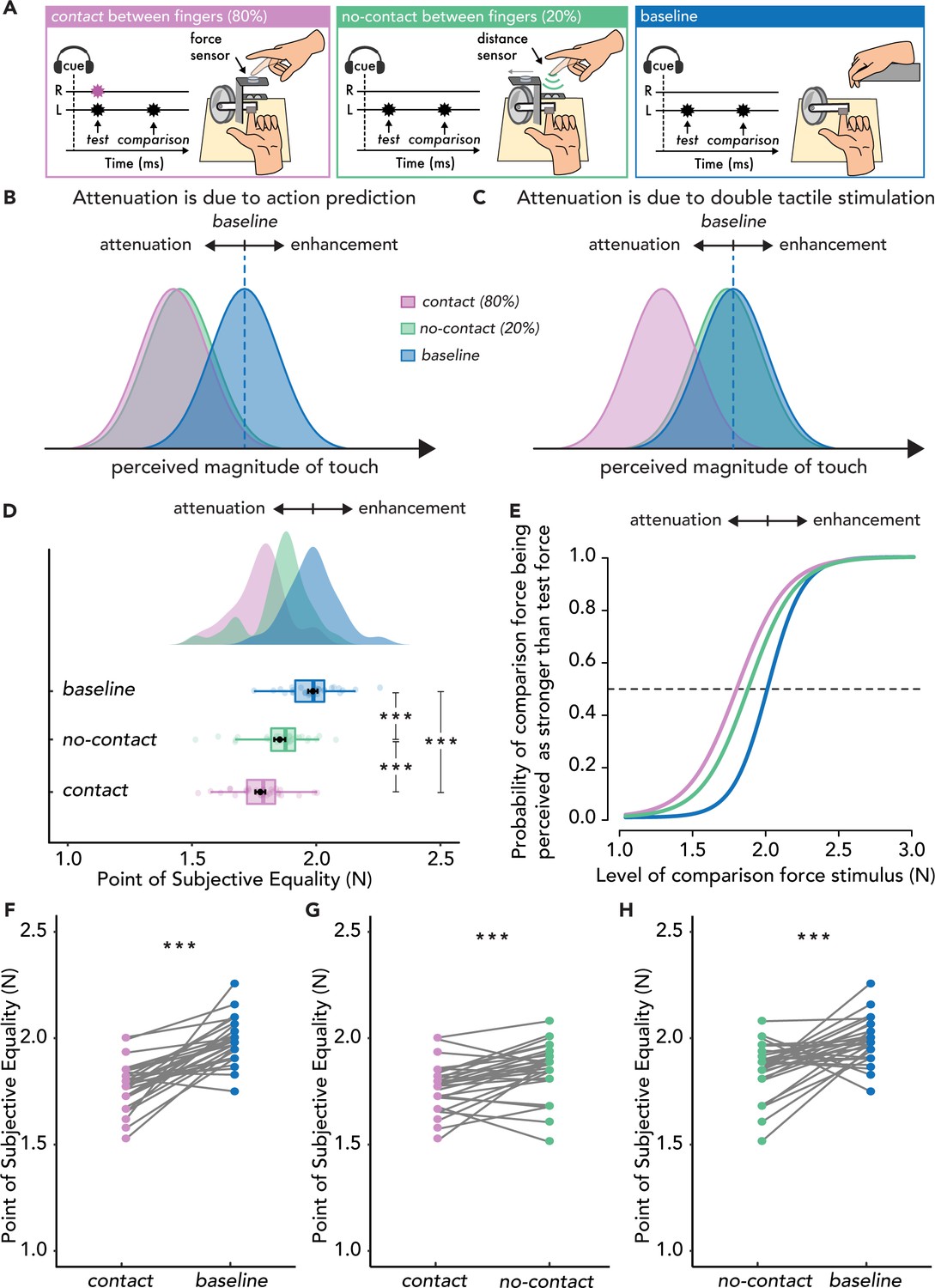
Experimental methods, hypotheses, and results of Experiment 3 (n=30).
(A) The contact (magenta) and no-contact (green) conditions were identical to those of Experiment 1, with the only difference being their relative proportion (contact trials 80%, no-contact trials 20%). In the no-contact trials, the force sensor was automatically retracted, unbeknownst to the participant, revealing the distance sensor placed below. The baseline condition (blue) was identical to that of Experiments 1 and 2. (B) Hypotheses based on the attenuation model. If attenuation is due to action prediction, then the perceived magnitude of touch should be reduced in both the contact (magenta) and no-contact conditions (green) compared to the baseline (blue). (C) Hypotheses based on the enhancement model. If attenuation effects are driven by simultaneous touch on the active hand, then the perceived magnitude of touch should be reduced only in the contact condition (magenta) compared to the baseline (blue) and not in the no-contact condition (green) which should be similar to the baseline (blue). (D–H) Data are color-coded per condition. (D) Box plots show the median and interquartile ranges for the point of subjective equality (PSE) values per condition, black circles and error bars show the mean PSE± SEM, and the raincloud plots show the individual PSE values and their distributions. (E) Group psychometric functions for each condition. The leftward shift of the curves in the contact and no-contact conditions indicates attenuated PSE values compared to the baseline. (F) Line plots for individual participant PSE values illustrate significantly lower PSEs for the contact versus baseline condition, (G) significantly lower PSEs in the contact versus no-contact condition and (H) significantly lower PSEs in the no-contact versus baseline. (***p<0.001).
The results showed a robust attenuation in both contact and no-contact trials with respect to the baseline condition, regardless of whether contact was made (Figure 3D, E and F & H): the PSEs were significantly lower in the contact condition than in the baseline condition (t(29) = 8.06, p<0.001, d = 1.47, CI95 = [0.16, 0.27]) and lower in the no-contact condition than in the baseline condition (t(29) = 4.45, p<0.001, d = 0.81, CI95 = [0.07, 0.20]). The magnitude of attenuation was larger in the contact condition than in the no-contact condition, with significantly lower PSEs in the contact condition than in the no-contact condition (t(29) = 3.94, p<0.001, d = 0.72, CI95 = [0.04, 0.12]) (Figure 3G). The difference in the magnitude of attenuation was modest (≅ 35%). These findings indicate that the perceived intensity of touch on the passive left hand was significantly attenuated both when the active hand received touch or not, thereby ruling out the possibility that attenuation is merely due to simultaneous tactile stimulation (Press et al., 2023; Thomas et al., 2022).
The results of Experiment 3 emphasize the predictive nature of somatosensory attenuation, which is observed only when the sensorimotor context allows the formation of such predictions. In Experiments 1 and 2 attenuation was only observed in the contact conditions, whereas attenuation was also observed in the no-contact condition of Experiment 3, where participants expected that they would make contact (i.e. the sensorimotor context was conducive to predicting self-touch). Thus, larger differences between the PSEs in the contact and no-contact conditions were observed for Experiments 1 and 2 compared to Experiment 3. To further strengthen our interpretation, we performed an ANOVA on the difference in the PSEs between the contact and no-contact conditions across all three experiments. The ANOVA revealed a significant main effect of Experiment (F(2, 87) = 8.05, p<0.001, = 0.156), with Bonferroni corrected post hoc comparisons indicating significant differences between Experiments 1 and 3 (t(58) = 3.10, p=0.008, d = 0.80, CI95 = [0.03, 0.23]) as well as Experiments 2 and 3 (t(58) = 3.76, p<0.001, d = 0.97, CI95 = [0.06, 0.26]), but no significant differences between Experiments 1 and 2 (t(58) = –0.66, p=1.000, d = –0.17, CI95 = [–0.13, 0.07], BF01 = 3.308). Similarly, an ANOVA on the difference in the PSEs between the baseline and no-contact conditions across all three experiments revealed a significant main effect of Experiment (F(2, 87) = 7.59, p<0.001, = 0.149). Bonferroni corrected post hoc comparisons indicated significant differences between Experiments 1 and 3 (t(58) = –3.76, p<0.001, d = –0.97, CI95 = [-0.27,–0.06]) and between Experiments 2 and 3 (t(58) = –2.76, p=0.021, d = –0.71, CI95 = [–0.22,–0.02]), but no significant difference between Experiments 1 and 2 (t(58) = –0.99, p=0.963, d = –0.26, CI95 = [–0.16, 0.06], BF01 = 2.537).
Thus, no-contact trials elicited attenuated perceptions only when the sensorimotor context allowed for a prediction of self-touch (Experiment 3) and not when the action was only arbitrarily associated with the touch (Experiments 1 and 2).
Regarding the JNDs, the values were significantly lower in the baseline condition than in the contact condition (t(29) = –3.61, p=0.001, d = –0.66, CI95 = [–0.09,–0.02]) and the no-contact condition (t(29) = –2.77, p=0.010, d = –0.51, CI95 = [–0.08,–0.01]), but there were no significant differences in the JNDs between the contact and no-contact conditions (t(29) = –0.65, p=0.521, d = –0.12, CI95 = [–0.02, 0.04], BF01 = 4.233). Thus, in contrast to the PSEs, there were no significant differences in the JNDs between the contact and no-contact conditions in any of the three experiments, demonstrating that the two conditions yielded the same discrimination capacity.
Kinematic results
To ensure that the movements in both the contact and no-contact conditions were comparable, participants were trained to make the same movement with their right index finger between two visual markers positioned 5 cm apart in Experiments 1 and 2. This training was not required for Experiment 3, in which participants were blindfolded and unaware that the distance sensor had been retracted on no-contact trials.
Given that the right index finger movement ends on impact with the force sensor in the contact condition but not in the no-contact condition, the kinematics of the finger movements are likely to differ between conditions. The active finger was slightly closer to the passive finger at the time of the test force in the contact compared to the no-contact condition in Experiments 1 and 2 (by less than 5 mm). Specifically, in Experiment 1, the position of the right index finger at the time of the test force was marginally closer to the left index finger in the contact condition than in the no-contact condition (mean difference = 2.32 mm, SEM = 1.14: t(29) = –2.04, p=0.051, d = 0.37, CI95 = [–0.74, 0.01], BF01 = 0.845). In Experiment 2, the position of the right index finger at the time of the test force was slightly closer to the left index finger in the contact condition than in the no-contact condition (mean difference = 4 mm, SEM = 1.8: t(29) = –2.29, p=0.030, d = 0.42, CI95 = [–0.79,–0.04]). In Experiment 3, participants were unaware that they would miss the force sensor in the no-contact trials, so with no force sensor to stop the movement, their right index finger continued further downwards. Here, the position of the right index finger at the time of the test force was closer to the left index finger in the no-contact condition than in the contact condition (mean difference = 25.7 mm, SEM = 2.0: t(29) = 12.79, p<0.001, d = 2.34, CI95 = [1.63, 3.03]). Critically, the closer position of the right index finger to the left index finger in the no-contact trials compared to the contact trials cannot explain the attenuation findings in the no-contact condition since the attenuation in the contact trials was stronger compared to that in the no-contact trials (Figure 3—figure supplement 2).
It could be argued that our differences between the contact and no-contact conditions across experiments are driven by differences in the timing of the two setups. This explanation is unlikely since we observed robust attenuation in the no-contact condition of Experiment 3 but not in Experiments 1 and 2 while the setups remained the same. To further control that the attenuation observed in the no-contact condition of Experiment 3 but not in the no-contact conditions of Experiments 1 and 2 was not driven by timing differences in the participants’ movement between experiments, we calculated the time the test force was delivered in the contact and no-contact conditions with respect to their movement endpoint (i.e. the force sensor press in the contact condition and the lowest vertical position in the no-contact condition). On average, the test force was applied 24 ms earlier in the no-contact compared to the contact condition in Experiment 1 (SEM = 4.20), 11 ms earlier in the no-contact compared to the contact condition in Experiment 2 (SEM = 4.53) and 15 ms earlier in the no-contact compared to the contact condition in Experiment 3 (SEM = 2.67). An ANOVA revealed a significant main effect of Experiment (F(2, 87) = 3.32, p=0.041, = 0.071), with Bonferroni corrected post hoc comparisons indicating significant differences between Experiment 1 and Experiment 2 (t(58) = 2.50, p=0.043, d = 0.65, CI95 = [0.62, 26.83], BF01 = 0.500). Importantly, there were no significant differences between neither Experiments 1 and 3 (t(58) = 1.80, p=0.226, d = 0.47, CI95 = [–3.21, 22.99], BF01 = 0.744), nor Experiments 2 and 3 (t(58) = –0.69, p=1.000, d = –0.18, CI95 = [–16.93, 9.27], BF01 = 3.051) demonstrating that the attenuation observed in Experiment 3 cannot be attributed to time differences between the participants’ movement and the received touch.
Discussion
In the present study, two opposing hypotheses regarding how action influences somatosensory perception were contrasted: the attenuation model and the enhancement model. Our findings demonstrate that action does not enhance (Experiments 1 and 2) but attenuates the predicted touch (Experiment 3) compared to identical touch in the absence of action. Before discussing the findings, it is important to emphasize that to draw conclusions about whether perception is attenuated or enhanced in an experimental condition including action, it is necessary to include a baseline condition without action. Only comparing conditions that include action prevents differentiating a genuine effect of enhancement (or attenuation) from an effect of reduced attenuation (or enhancement) in one of the two conditions. This also applies to experimental manipulations that contrast predicted with unpredicted somatosensory stimuli during action (see Thomas et al., 2022 for such comparisons). To this end, a baseline condition of pure somatosensory exafference (i.e. externally generated touch) was included in all the present experiments that allowed us to distinguish the direction of the effects.
The results of Experiment 1 showed a robust attenuation of the touch applied to the passive left hand when the two hands made contact (contact condition), but neither attenuation nor enhancement was caused by the mere movement of the right hand (no-contact condition). In contrast, the perceived intensity of touch in the no-contact condition was similar to that of the baseline (i.e. externally generated touch). These findings are in line with the results of Bays et al., 2006, who found no change in the participants’ somatosensory perception when the two hands did not make contact, but do not replicate those of Thomas et al., 2022, who observed enhancement of touch in a no-contact condition. Experiment 2 further investigated the source of the previously reported enhancement effects and showed that they are in fact driven by the reference condition used: enhancement was observed only relative to a condition in which the touch was applied rapidly following a cue to inhibit the movement (i.e. a ‘do not move’ cue), as in previous research (Thomas et al., 2022), but not relative to an externally generated touch condition (our baseline). In support of this claim, Experiment 2 showed that a cue to inhibit the movement results in a significant reduction in the perceived intensity of the imperative stimulus (i.e. touch on the passive hand) compared to baseline perception. This is in line with previous evidence showing reduced amplitudes of somatosensory evoked potentials for tactile stimuli presented shortly following a cue to inhibit a movement (Hoshiyama and Sheean, 1998), reduced perceived amplitude for tactile stimuli under the mere expectation to move (Voss et al., 2008; Voss et al., 2006), and reduced encoding of sensory stimuli following motor inhibition (Chiu and Egner, 2015b; Yebra et al., 2019). Therefore, rather than participants’ perception being enhanced in the no-contact condition, it is a reduction of the perceived intensity following the cue to inhibit the movement that leads to an apparent enhancement. In contrast, by including a novel externally generated touch condition as a baseline that involved neither motor planning nor response inhibition, it became clear that there were no enhancement effects.
Experiment 3 demonstrated that action attenuates the predicted touch even if the active hand does not receive simultaneous tactile stimulation with the passive hand. When participants simply moved their right hand to trigger the touch on their left hand (no-contact trials in Experiments 1 and 2), no change in their somatosensory perception was found. This suggests that an arbitrary mapping between the movement of the right hand and the delivery of touch on the left hand is insufficient to elicit attenuation. In contrast, when participants expected that they would touch their own hand (Experiment 3), significant attenuation was observed, even when the active hand unexpectedly missed the contact (no-contact trials, 20%). This pattern of results can be explained by the attenuation model, which suggests that the self-generation of a stimulus is in itself not sufficient for predictive attenuation, but that the action and its predicted sensory feedback are consistent with touching one’s own body (Bays and Wolpert, 2008). Thus, a sensorimotor context that closely resembles tapping directly on the left index finger with the right index finger (self-touch) was critical for predictive attenuation (Bays et al., 2006). The findings of Experiment 3 also contradict the suggestion that attenuation on a passive hand reflects a nonpredictive gating of tactile input during movement of the active hand (Press et al., 2023; Thomas et al., 2022). Indeed, several earlier studies showed gating effects only on the moving limb (movement effector) and not on the passive limb (Chapman et al., 1987; Cohen and Starr, 1987; Colino et al., 2014; Papakostopoulos et al., 1975; Pertovaara et al., 1992; Rushton et al., 1981), and we have also recently shown that experimental paradigms identical to the one used in the present study produce attenuation effects but not gating effects on the passive hand (Kilteni and Ehrsson, 2022). Overall, the results suggest that attenuation effects are driven by action prediction and not the simultaneous tactile stimulation of the two hands. From an ecological point of view, in every self-touch behavior, we necessarily receive somatosensory input on the active hand and the body part that passively receives touch (‘touchant-touché’ [Schütz-Bosbach et al., 2009]), and it is within these sensorimotor contexts that the brain forms predictions about the somatosensory consequences on multiple body parts (Bays and Wolpert, 2008).
Interestingly, the magnitude of attenuation was greater in the contact compared to the no-contact condition by approximately 35% in Experiment 3. This could suggest that the attenuation effect observed in the contact condition arises from multiple sources, one being the motor-based prediction and another source being the simultaneous touch on the active hand. However, this interpretation is inconsistent with the findings that somatosensory attenuation has been previously observed in the absence of simultaneous tactile stimulation, for example, when imagining but not executing the right hand movement (Kilteni et al., 2018) or just before the hands make contact (Bays et al., 2005). Similarly, no somatosensory attenuation is observed in the absence of action prediction, even if the two hands receive simultaneous tactile stimulation; for example, the passive displacement of the right hand towards the left hand that is accompanied by a double touch (Kilteni et al., 2020) or the mere delivery of simultaneous tactile stimulation on both hands (Bays et al., 2005) does not produce attenuation. Instead, our finding that the magnitude of attenuation was greater in the contact than the no-contact condition in Experiment 3 may instead reflect a decrease of attenuation in the no-contact trials, due to the unexpected omission of contact influencing the perceived magnitude of touch in a postdictive manner. Specifically, the unexpected omission of contact in the minority of trials (20%) could be seen as a form of stimulus omission akin to so-called ‘silent oddballs’ that are known to generate prediction errors (Busse and Woldorff, 2003; Karamürsel and Bullock, 2000; SanMiguel et al., 2013a; SanMiguel et al., 2013b). That is, although participants clearly attenuated the touch predicted by their movement in the no-contact trials, their expectation of contact was necessarily violated. In one of our previous studies (Kilteni et al., 2019) using the same force discrimination task as in the present study we showed in three experiments that participants experienced the touch to be stronger (i.e. less attenuation) and more ticklish when triggering a prediction error (<17% frequency). Similarly, research outside the domain of action (Kusnir et al., 2020) has found that perception is improved for tactile events delivered at unpredictable compared to predictable temporal onsets, also suggesting that tactile perception is facilitated in the presence of prediction error. In line with these previous results, the decreased attenuation we observed in the no-contact condition could be attributed to the prediction error. Additionally, this violation could be considered a novel event given its infrequency. Novel events can have several consequences for cognition, including transient enhancements of perception (Schomaker and Meeter, 2012), facilitated encoding of information into working memory (Mayer et al., 2011) as well as changes in the allocation of attentional resources in a postdictive manner (for a review of the effects of novelty on cognition see Schomaker and Meeter, 2015).
Some authors have criticized the comparison of perceived intensity of sensory stimuli in self-generated and externally generated conditions (Press et al., 2020; Yon et al., 2018; Yon et al., 2021). A frequent criticism is that tactile input may be ‘predicted’ in self-generated conditions through action and ‘unpredicted’ in externally generated (passive) conditions that do not involve action. This concern can be ruled out since in all three experiments, the stimulus in the baseline (i.e. externally generated touch) condition was delivered at a fixed timepoint (800 ms after the cue); therefore, participants could predict it in the absence of motor-based predictions. Second, it has been argued that somatosensory attenuation findings may be driven by dual-task requirements (Press et al., 2020) present only in self-generated conditions that could increase the working memory load or result in a shift of attention towards the active hand. A similar criticism has been suggested in which the mere movement of the active hand in self-generated conditions can result in a nonpredictive gating of tactile input. However, these explanations can also be ruled out, given that dual-task requirements and movement of the active hand were present in the contact and no-contact conditions of all three experiments without concomitant attenuation effects. Indeed, if dual-task requirements, or the mere presence of movement during the perceptual task, were sufficient to cause attenuation effects, then attenuation would have been observed in the no-contact conditions of Experiments 1 and 2. Furthermore, if the mere simultaneous tactile stimulation on the active hand was responsible for the attenuation effect, we would not have observed attenuation in the no-contact condition of Experiment 3. Finally, alternative explanations based on differences in other psychophysical parameters, movement kinematics, or timings between experiments can also be ruled out.
How can the findings that action prediction attenuates touch be reconciled with those showing that expectations outside the domain of action improve sensory perception (Press et al., 2020)? While it is difficult to directly compare these lines of research because of differences in the sensory modality investigated, the task designs used, and the perceptual measures employed, we speculate that there are numerous possible reasons why action prediction may not have the same effect on perception as prediction mechanisms outside the domain of action. First, research on action-based predictions concerns ubiquitous associations between actions and their sensory consequences that we are continually exposed to throughout the lifespan. For example, we are constantly exposed to associations between our motor behaviors and their tactile consequences during self-touch, even as early as 13 weeks in utero (Kurjak et al., 2003). In contrast, research on sensory expectations outside the domain of action primarily concerns arbitrary associations between stimuli that are typically learned only during the time course of a given task. It is, therefore, conceivable that separable mechanisms may operate to predict action effects versus stimulus-stimulus associations (see Dogge et al., 2019a; Dogge et al., 2019b for discussion). Second, higher-level expectations, such as explicit prior knowledge that a specific sensory event is likely, might not operate in the same way as lower-level action predictions; for example, it has been proposed that action-based predictions inhibit expected stimuli, while sensory expectations potentiate the expected sensory inputs (de Lange et al., 2018). Most importantly, from a theoretical perspective, attenuating the predicted sensory consequences of actions does not necessarily mean that the brain forms inaccurate representations of the world but instead indicates a flexible strategy that prioritizes more informative externally generated events.
It is important to emphasize that this study investigated the perception of self-generated touch during an action directed toward one’s own body. Thus, perception was assessed on a passive body part that was the target of a movement of an active body part (i.e. moving the right hand to touch the static left hand). This focus was chosen because extensive previous research has shown deteriorated performance when detecting or discriminating touch (both self-generated and externally generated) delivered on a moving limb, a phenomenon referred to as movement-related gating (Chapman et al., 1996; Chapman et al., 1987; Voudouris and Fiehler, 2021; Voudouris and Fiehler, 2017; Williams et al., 1998). By assessing somatosensory perception on a static limb, we were able to isolate the effect of action prediction on somatosensory perception from other perceptual phenomena related to the movement per se including movement-related gating (Chapman et al., 1987; Cohen and Starr, 1987; Colino et al., 2014; Kilteni and Ehrsson, 2022; Papakostopoulos et al., 1975; Pertovaara et al., 1992; Rushton et al., 1981). However, it is interesting to note that during the active exploration of objects through touch, referred to as active touch, self-generated touch also occurs, albeit only on the moving limb. Studies on active touch have observed improved perception of object features, such as texture or edge orientation when actively exploring an object (Morley et al., 1983; Pruszynski et al., 2018) compared to passive exposure (Bensmaia et al., 2008; Peters et al., 2015). At first, the improved perception observed during active touch might appear paradoxical to observations of movement-related gating and predictive attenuation effects: if anything, active touch should result in worse perception than passive touch. However, it has been shown that humans optimize their exploratory movements to minimize these effects and maximize the quality of their sensory feedback. For example, it is known that the degree of gating decreases as movement speed slows (Angel and Malenka, 1982; Chapman et al., 1996; Schmidt et al., 1990) and it has been proposed that participants optimize the speed of movement during active touch to avoid high speeds associated with tactile gating (Cybulska-Klosowicz et al., 2011). Similarly, when exploring objects through active touch, weaker forces are typically applied (e.g. <1 N [Olczak et al., 2018; Tanaka et al., 2014]) that elicit less attenuation than stronger forces, as shown in the classic force-matching task (Shergill et al., 2003). Thus, differences in the task context might primarily influence how one moves (e.g. the act of swatting a fly from one’s arm versus feeling for a bump on one’s skin), which can have disparate ensuing effects on perception depending on the movement kinematics.
Debates between researchers supporting attenuation or enhancement are useful and fruitful for scientific dialogue and advancement. The present study revisited recent findings on somatosensory enhancement during action and showed that when the requirement to inhibit the action is removed from the baseline, action predictions do not enhance but attenuate the received somatosensory input. Our results are in strong alignment with animal studies showing that action attenuates the predicted sensory consequences (for reviews, see [Brooks and Cullen, 2019; Crapse and Sommer, 2008; Cullen, 2004; Schneider and Mooney, 2018; Straka et al., 2018]). For example, crickets and mice suppress auditory reafferent signals but maintain their sensitivity to external sounds (Audette et al., 2022; Poulet and Hedwig, 2003; Poulet and Hedwig, 2006; Schneider et al., 2018), the weakly electric fish attenuates its electrosensory reafference to respond to externally generated electrical signals (Cullen, 2004; Sawtell, 2017), and primates attenuate their vestibular reafference and activate vestibular-related reflexes only when the vestibular input is exafferent (Brooks et al., 2015; Cullen, 2012; Roy and Cullen, 2004). To this end, the results of the present study prompt a reappraisal of recent experimental findings upon which theoretical frameworks proposing a perceptual enhancement by action prediction are based.
Methods
Participants
Thirty naive adults participated in Experiment 1 (18 females, aged 18–36, 28 right-handed and 2 ambidextrous), thirty naive adults participated in Experiment 2 (17 females, aged 20–37, 29 right-handed and 1 left-handed) and thirty naive adults participated in Experiment 3 (12 females, aged 21–40, 28 right-handed, 1 ambidextrous, 1 left-handed). Current or history of psychological or neurological conditions, as well as the use of any psychoactive drugs or medication to treat such conditions, were criteria for exclusion. The sample size of each Experiment was decided prior to data collection based on our previous studies using the same task (Kilteni et al., 2019; Kilteni and Ehrsson, 2022). Handedness was assessed using the Edinburgh Handedness Inventory (Oldfield, 1971). All experiments were approved by the Swedish Ethical Review Authority (registration no. 2021–03790). Participants provided written informed consent and were compensated for their time.
Pre-registration of experiments
The method and analysis plan for each experiment was pre-registered on the Open Science Framework (OSF) prior to data collection (Experiment 1 https://osf.io/9jkqt, Experiment 2 https://osf.io/hs8au, Experiment 3 https://osf.io/gkwu7).
Modifications/additions to the pre-registered plan
The only modification from the pre-registered plan was a reduction in the number of trials administered in Experiment 3. Following initial piloting, it was deemed appropriate to reduce the overall length of the task by including fewer trials (see Methods details). Three additional ANOVAs were also conducted to compare the results across the three experiments and the kinematics across experiments.
Experimental setup
Participants sat comfortably on a chair with their arms placed on a table. The left hand rested palm up, with the index finger placed on a molded support. On each trial, a motor (Maxon EC Motor EC 90 flat; Switzerland) delivered two forces (the test force and the comparison force) on the pulp of the left index finger through a cylindrical probe (25 mm height) with a flat aluminum surface (20 mm diameter) attached to a lever on the motor. A force sensor (FSG15N1A, Honeywell Inc; diameter, 5 mm; minimum resolution, 0.01 N; response time, 1 ms; measurement range, 0–15 N) within the probe recorded the forces applied on the left index finger. Following the presentation of the two forces, participants were asked to verbally report which of the two forces felt stronger – the first or the second. A second identical force sensor within an identical cylindrical probe (‘active force sensor’) was placed on top of, but not in contact with, the probe of the left index finger.
Force-discrimination task
Participants judged the intensity of a test force and a comparison force (100 ms duration each) separated by a random interval between 800 ms and 1200 ms in a two-alternative forced choice (2AFC) task. The intensity of the test force was 2 N, while the intensity of the comparison force was systematically varied among seven force levels (1, 1.5, 1.75, 2, 2.25, 2.5, or 3 N). In all the conditions, the forces were delivered by the same motor, to precisely control their magnitude. However, the source of the force was manipulated across conditions such that the force was triggered by the participants’ contact with a force sensor (contact condition), their finger movement (no-contact condition), or automatically by the stimulus computer (baseline condition).
Experimental design and procedures
Experiment 1
There were three conditions in Experiment 1 presented in three blocks separately: the baseline (i.e. externally generated touch) condition, the contact condition, and the no-contact condition.
In the baseline condition, participants did not move their limbs but passively received the test and the comparison force on the left index finger. This baseline condition was used to assess the participants’ somatosensory perception in the absence of any movement (Bays et al., 2005; Kilteni et al., 2020; Kilteni et al., 2019). Each trial began with an auditory cue (100 ms duration, 997 Hz) followed by the test force delivered to the participant’s left index finger 800 ms after the cue by the motor. The comparison force was then delivered at a random interval between 800 ms and 1200 ms after the test force.
In the contact condition, participants started each trial by holding their right index finger approximately 10cm above their left index finger. The start position was marked with a visual marker placed next to their right index finger while the final position was the probe of the active force sensor. The same auditory cue was presented as in the baseline condition, but participants now moved their right index finger downwards toward their left index finger and actively tapped on the active force sensor placed on top of, but not in contact with, the probe. The participant’s active tap on the force sensor triggered the motor to apply the test force on their left index finger (threshold 0.2N). The tap of their right index finger triggered the test force on their left index finger with an intrinsic delay of 36 ms (Figure 3—figure supplement 3). The comparison force was then delivered at a random interval between 800 ms and 1200 ms after the test force. As in our previous studies (Asimakidou et al., 2022; Kilteni et al., 2021; Kilteni and Ehrsson, 2022), participants were asked to tap, neither too weakly nor too strongly, with their right index finger, ‘as if tapping the screen of their smartphone’. This instruction was provided to ensure that the relationship between the force they applied with their right index finger on the force sensor and the force they received on their left index finger by the motor (2N) remained approximately constant throughout the experiment, thereby establishing perceived causality (Bays and Wolpert, 2008; Kilteni, 2023). Indeed, across all three experiments, we observed low variability in the exerted forces (Figure 3—figure supplement 3D-F ) (± 0.13N SEM in Experiment 1, ± 0.12N SEM in Experiment 2 and ± 0.11N SEM in Experiment 3) establishing an approximately constant gain between the exerted and the applied forces. Critically, it has been previously shown that the magnitude of attenuation remains unaffected by halving or doubling the gain between the force applied by the active finger and the force delivered on the passive hand as long as the gain remains constant (Bays and Wolpert, 2008). This should not be surprising given that when one finger transmits a force through an object to another finger, the resulting force depends also on the object’s properties (e.g. shape, material, contact area) and the angle at which the finger contacts the object.
In the no-contact condition, the participants started each trial by holding their right index finger 10cm above their left index finger, identically to the contact condition. For this block, the active force sensor was removed and replaced by a distance sensor that detected the position of the right index finger. The distance sensor was placed on top of, but not in contact with, the probe. Following the same auditory cue, participants moved their right index finger towards their left index finger. To restrict the participants’ right index finger movement within similar movement ranges as in the contact condition and avoid contact with the distance sensor placed underneath, a second visual marker indicated the final position of the right index finger above the distance sensor (5cm from the marker indicating the initial position). The distance sensor was connected to an Arduino microcontroller that controlled a servo motor. The servo motor and the force sensor were placed 2 meters away from the participants’ hands and hidden from view. Once the distance sensor detected that the position of the right index finger became smaller than a preset threshold, it triggered the servo motor that hit the force sensor and triggered the test force. We accounted for the additional delay of the distance sensor by setting the position threshold for the distance sensor slightly higher than the lowest position the participants were asked to reach with their right index finger. Therefore, the test force would be delivered at a similar timing to the participants’ right index finger endpoint between the contact and no-contact conditions. This position threshold was set based on significant pilot testing prior to all experiments. Indeed, there was a minimal time difference between the two setups across experiments (17 ms average delay) with the no-contact condition leading, rather than lagging the contact one (see Results). The comparison force was delivered at a random interval between 800 ms and 1200 ms after the test force. Therefore, in the no-contact condition, there was no touch on the right index finger simultaneously with the test force on the left index finger.
Before the experiment, participants were trained to make similar movements with their right index finger in the contact and no-contact conditions, emphasis was placed on restricting their right index finger movements to between the two visual markers in the no-contact condition. The 3D position of the right index finger was recorded using a Polhemus Liberty tracker (240Hz). Kinematic information was used to compare the movements between the contact and no-contact conditions and to reject any trials in which the participant did not trigger the test stimulus with their movement, or trials in which the participant did not move as instructed in the training (see below). Participants were administered white noise through headphones to mask any sounds made by the motor to serve as a cue for the task. The loudness of the white noise was adjusted such that participants could hear the auditory tones of the trial. In all conditions the participants’ left index finger as well as the motor that delivered the force to their left index finger, was occluded from vision. This was done to prevent participants using any visual cues to discriminate between the two forces.
Each block consisted of 70 trials resulting in 210 trials per participant. Thus, the proportion of contact and no-contact trials was the same (50–50%). The order of the conditions was fully counterbalanced across participants.
Experiment 2
The 2-AFC task was identical to that of Experiment 1. In Experiment 2, there were five conditions in total: the baseline condition, the contact condition, the no-contact condition, and two additional NOGO conditions (NOGO contact and NOGO no-contact). Trials of the NOGO conditions were pseudo-randomly intermixed with trials of the contact and no-contact conditions, respectively (GO trials). The five conditions were presented in three separate blocks: the baseline, contact (GO and NOGO trials), and no-contact (GO and NOGO trials) blocks.
In the contact block, 50% of trials began with an auditory ‘GO’ cue (100 ms duration high tone of 2458Hz) instructing participants to tap the active force sensor to trigger the test force (identically to the contact condition of Experiment 1). On the remaining 50% of trials an auditory ‘NOGO’ cue (100 ms duration low tone of 222Hz) instructed participants to withhold their movement and the test force was then delivered 800 ms following the NOGO cue. The no-contact block was identical to the contact block, except that the test force was triggered by the position of the right index finger without contact with the force sensor (identically to the no-contact condition of Experiment 1). The cue tone in the baseline block was the same as the cue tone in the NOGO trials (100 ms duration low tone of 222Hz).
Therefore, this design replicated the experimental design of Thomas et al., 2022 with the additional inclusion of the baseline (i.e. externally generated touch) condition. As in Experiment 1, we recorded the 3D position of the right index finger, and the registered kinematic information was used to compare the movements between the contact and no-contact conditions and reject any trials in which the participants did not trigger the test stimulus with their movement, trials in which the participants did not move as instructed in the training (see below), and trials in which the participants moved while instructed not to do so by the auditory cues (NOGO trials). As in Experiment 1, participants were administered white noise and the loudness of the white noise was adjusted such that participants could clearly hear the GO and NOGO auditory cues of the trial.
The baseline block consisted of 70 trials, and the contact and no-contact blocks consisted of 70 GO trials and 70 NOGO trials each, resulting in 350 trials in total per participant. The order of the blocks was fully counterbalanced across participants.
Experiment 3
The 2-AFC task and the experimental conditions (baseline, contact, and no-contact) were identical to those of Experiment 1, except for the following. Contact trials (80%) were now pseudo-randomly intermixed with no-contact trials (20%) within the same block. The force sensor was now attached to a plastic platform that could be automatically retracted by a servo motor, depending on the trial type. Upon retraction, a distance sensor placed underneath the platform was revealed. In the contact trials, participants tapped the force sensor to trigger the test force identically to Experiments 1 and 2. In the no-contact trials, the platform was automatically retracted before trial onset, unbeknownst to the participants. This led participants to unexpectedly miss the active force sensor and instead trigger the test force only by the position of their right index finger. In all conditions, the participants’ vision was occluded with a blindfold, and white noise was administered via headphones to prevent any visual or auditory cues indicating that the force sensor had been retracted in no-contact trials. Note that the blindfold was used for all conditions of Experiment 3 to control for any effects of lack of vision between conditions and allow comparisons between conditions (e.g. baseline minus no-contact, no-contact minus contact) with those of Experiments 1 and 2. The baseline block was identical to that of Experiments 1 and 2, except for the use of a blindfold.
In Experiment 3, the number of trials was reduced from 70 to 56 to shorten the total experiment time to less than 90 min, similar to Experiment 2. Thus, there were 56 no-contact trials (20%) and 224 contact trials (80%). Similarly, there were 56 trials in the baseline condition. This resulted in 336 trials per participant. The order of the two blocks was fully counterbalanced across participants.
Preprocessing of psychophysical trials
Experiment 1
Experiment 1 included 6300 trials in total (30 participants * 70 trials * 3 conditions). Twenty-nine (29) trials were excluded (0.5%) because of 1 missing response, four trials in which the force was not applied correctly (1.85 N<test force <2.15 N), and 24 trials because the test force was not triggered when moving towards the distance sensor. The kinematic analysis also excluded all trials excluded by the psychophysical fits.
Experiment 2
Experiment 2 included 10,500 trials in total (30 participants * 70 trials * 5 conditions). One-hundred fourteen (114) trials were excluded (1.1%) because of two missing responses, six trials in which the force was not applied correctly (1.85 N<test force <2.15 N), 44 trials because the test force was not triggered when moving towards the distance sensor and 62 trials because the finger moved on a NOGO trial. All trials excluded by the psychophysical fits were also excluded in the kinematic analysis.
Experiment 3
Experiment 3 included 10,080 trials in total (30 participants * (224 contact trials + 56 no-contact trials + 56 baseline trials)). Two-hundred twenty-four (29) trials were excluded (2.42%) because of 29 missing responses, 86 trials in which the force was not applied correctly (1.85 N<test force <2.15 N), 81 because the test force was not triggered when moving towards the distance sensor and 48 because the finger contacted the distance sensor on no-contact trials. The kinematic analysis also excluded all trials excluded by the psychophysical fits.
Preprocessing of kinematic recordings
All kinematic trials were co-registered with the force trials through Transistor-Transistor Logic (TTL) signals sent by the motor to both file outputs. The kinematic recordings were corrected for any distortion in the Polhemus sensor from the force sensor and distance sensor based on measurements made with and without the force/distance sensors within the same space. Exclusion of trials based on the kinematics was done by assessing whether the test force was delivered after the position of the active finger reached its minimum value on the vertical plane. Trials in which the force was delivered after the minimum value had been reached were rejected. For experiment 2, trials in which the active finger moved more than 1 cm following a NOGO cue were also rejected.
Fitting of psychophysical responses
For each experiment and each condition, the participant’s responses were fitted with a generalized linear model using a logit link function (Equation 1):
(1)
We extracted two parameters of interest: the Point of Subjective Equality (PSE) (), which represents the intensity at which the test force felt as strong as the comparison force (p=0.5) and quantifies the perceived intensity, and the Just Noticeable Difference (JND) (), which reflects the participants’ discrimination ability. Before fitting the responses, the values of the applied comparison forces were binned to the closest value with respect to their theoretical values (1, 1.5, 1.75, 2, 2.25, 2.5, or 3 N).
In all three experiments, for all participants and all conditions, the fitted logistic models were very good, with McFadden’s R-squared measures ranging between 0.602 and 0.979 (Figure 1—figure supplement 1, Figure 2—figure supplement 1, Figure 3—figure supplement 1).
Normality of data and statistical comparisons
We used R (R Development Core Team, 2021), JASP (Team, 2022) and MATLAB (2020b) to analyze our data. The normality of the PSE and the JND data distributions, as well as the kinematic information data distributions were assessed using Shapiro-Wilk tests. Depending on the data normality, pairwise comparisons between conditions were performed by using either a paired t-test or a Wilcoxon signed-rank test. We report 95% confidence intervals (CI95) for each statistical test. Effect sizes are reported as the Cohen’s d for t-tests or the matched rank biserial correlation rrb for the Wilcoxon signed-rank tests. In addition, a Bayesian factor analysis using default Cauchy priors with a scale of 0.707 was performed for all statistical tests that led to not statistically significant effects, to provide information about the level of support for the null hypothesis compared to the alternative hypothesis (BF01) based on the data. We interpret a Bayes factor from 1 to 3 as providing 'anecdotal' support for the null hypothesis, and a Bayes factor from 3 to 10 as providing 'moderate' support for the null hypothesis (Quintana and Williams, 2018; van Doorn et al., 2021). For the Analysis of Variance (ANOVA), homogeneity of variance was assessed using Levene’s test for Equality of Variances, which did not reach significance, and the Q-Q plot of the standardized residuals indicated approximately normally distributed residuals. Post-hoc tests were made using Bonferroni corrections. All tests were two-tailed.
Data availability
Data (PSEs and JNDs) are publicly available along with pre-registered analyses plans on the OSF (Experiment 1: https://osf.io/9jkqt, Experiment 2: https://osf.io/hs8au, Experiment 3: https://osf.io/pxy87).
-
Open Science FrameworkID 9jkqt. Attenuation versus enhancement of self-generated touch.
-
Open Science FrameworkID hs8au. Attenuation of self-generated touch and inhibition.
-
Open Science FrameworkID pxy87. Attenuation of self-generated touch and active-hand contact.
References
-
Velocity-dependent suppression of cutaneous sensitivity during movementExperimental Neurology 77:266–274.https://doi.org/10.1016/0014-4886(82)90244-8
-
Precise movement-based predictions in the mouse auditory cortexCurrent Biology 32:4925–4940.https://doi.org/10.1016/j.cub.2022.09.064
-
Predictive attenuation in the perception of touchSensorimotor Foundations of Higher Cognition 1:0001.https://doi.org/10.1093/acprof:oso/9780199231447.001.0001
-
The tactile perception of stimulus orientationSomatosensory & Motor Research 25:49–59.https://doi.org/10.1080/08990220701830662
-
Central cancellation of self-produced tickle sensationNature Neuroscience 1:635–640.https://doi.org/10.1038/2870
-
Predicting the consequences of our own actions: the role of sensorimotor context estimationThe Journal of Neuroscience 18:7511–7518.https://doi.org/10.1523/JNEUROSCI.18-18-07511.1998
-
Spatio-temporal prediction modulates the perception of self-produced stimuliJournal of Cognitive Neuroscience 11:551–559.https://doi.org/10.1162/089892999563607
-
Why can’t you tickle yourself?Neuroreport 11:R11–R16.https://doi.org/10.1097/00001756-200008030-00002
-
Predictive sensing: The role of motor signals in sensory processingBiol Psychiatry Cogn Neurosci Neuroimaging 4:842–850.https://doi.org/10.1016/j.bpsc.2019.06.003
-
Active inference, sensory attenuation and illusionsCognitive Processing 14:411–427.https://doi.org/10.1007/s10339-013-0571-3
-
Sensory perception during movement in manExperimental Brain Research 68:516–524.https://doi.org/10.1007/BF00249795
-
BookFactors influencing the perception of tactile stimuli during movementSomesthesis and the Neurobiology of the Somatosensory Cortex.https://doi.org/10.1007/978-3-0348-9016-8_25
-
Inhibition-induced forgetting: when more control leads to less memoryPsychological Science 26:27–38.https://doi.org/10.1177/0956797614553945
-
Inhibition-induced forgetting results from resource competition between response inhibition and memory encoding processesThe Journal of Neuroscience 35:11936–11945.https://doi.org/10.1523/JNEUROSCI.0519-15.2015
-
Tactile gating in a reaching and grasping taskPhysiological Reports 2:e00267.https://doi.org/10.1002/phy2.267
-
Hallucinations and Strong PriorsTrends in Cognitive Sciences 23:114–127.https://doi.org/10.1016/j.tics.2018.12.001
-
Predicting to perceive and learning when to learnTrends in Cognitive Sciences 24:259–260.https://doi.org/10.1016/j.tics.2019.12.005
-
Corollary discharge across the animal kingdomNature Reviews. Neuroscience 9:587–600.https://doi.org/10.1038/nrn2457
-
Sensory signals during active versus passive movementCurrent Opinion in Neurobiology 14:698–706.https://doi.org/10.1016/j.conb.2004.10.002
-
The vestibular system: multimodal integration and encoding of self-motion for motor controlTrends in Neurosciences 35:185–196.https://doi.org/10.1016/j.tins.2011.12.001
-
A critical speed for gating of tactile detection during voluntary movementExperimental Brain Research 210:291–301.https://doi.org/10.1007/s00221-011-2632-0
-
Widespread access to predictive models in the motor system: A short reviewJournal of Neural Engineering 2:S313–S319.https://doi.org/10.1088/1741-2560/2/3/S11
-
How do expectations shape perception?Trends in Cognitive Sciences 22:764–779.https://doi.org/10.1016/j.tics.2018.06.002
-
Moving forward: On the limits of motor-based forward modelsTrends in Cognitive Sciences 23:743–753.https://doi.org/10.1016/j.tics.2019.06.008
-
Explaining the symptoms of schizophrenia: abnormalities in the awareness of actionBrain Research. Brain Research Reviews 31:357–363.https://doi.org/10.1016/s0165-0173(99)00052-1
-
The neural basis of hallucinations and delusionsComptes Rendus Biologies 328:169–175.https://doi.org/10.1016/j.crvi.2004.10.012
-
The self in action: Lessons from delusions of controlConsciousness and Cognition 14:752–770.https://doi.org/10.1016/j.concog.2005.04.002
-
Can a problem with corollary discharge explain the symptoms of schizophrenia?Biological Psychiatry. Cognitive Neuroscience and Neuroimaging 4:768–769.https://doi.org/10.1016/j.bpsc.2019.07.003
-
One step ahead: The perceived kinematics of others’ actions are biased toward expected goalsJournal of Experimental Psychology. General 145:1–7.https://doi.org/10.1037/xge0000126
-
Human auditory fast and slow omitted stimulus potentials and steady-state responsesThe International Journal of Neuroscience 100:1–20.https://doi.org/10.3109/00207450008999674
-
No evidence for somatosensory attenuation during action observation of self-touchThe European Journal of Neuroscience 54:6422–6444.https://doi.org/10.1111/ejn.15436
-
BookMethods of Somatosensory AttenuationIn: Holmes NP, editors. Somatosensory Research Methods. Springer. pp. 35–53.https://doi.org/10.1007/978-1-0716-3068-6
-
The spatial specificity of sensory attenuation for self-touchConsciousness and Cognition 92:103135.https://doi.org/10.1016/j.concog.2021.103135
-
Fetal hand movements and facial expression in normal pregnancy studied by four-dimensional sonographyJournal of Perinatal Medicine 31:496–508.https://doi.org/10.1515/JPM.2003.076
-
When temporal certainty doesn’t helpJournal of Cognitive Neuroscience 32:315–325.https://doi.org/10.1162/jocn_a_01482
-
Internal models in biological controlAnnual Review of Control, Robotics, and Autonomous Systems 2:339–364.https://doi.org/10.1146/annurev-control-060117-105206
-
Methodological considerations for the force-matching taskBehavior Research Methods 55:2979–2988.https://doi.org/10.3758/s13428-022-01954-w
-
Tactile discrimination of gratingsExperimental Brain Research 49:291–299.https://doi.org/10.1007/BF00238588
-
Edge orientation perception during active touchJournal of Neurophysiology 120:2423–2429.https://doi.org/10.1152/jn.00280.2018
-
Lowered cutaneous sensitivity to nonpainful electrical stimulation during isometric exercise in humansExperimental Brain Research 89:447–452.https://doi.org/10.1007/BF00228261
-
Corollary discharge inhibition of ascending auditory neurons in the stridulating cricketThe Journal of Neuroscience 23:4717–4725.https://doi.org/10.1523/JNEUROSCI.23-11-04717.2003
-
Perceptual prediction: Rapidly making sense of a noisy worldCurrent Biology 29:R751–R753.https://doi.org/10.1016/j.cub.2019.06.054
-
The perceptual prediction paradoxTrends in Cognitive Sciences 24:13–24.https://doi.org/10.1016/j.tics.2019.11.003
-
Cancelling cancellation? Sensorimotor control, agency, and predictionNeuroscience and Biobehavioral Reviews 145:105012.https://doi.org/10.1016/j.neubiorev.2022.105012
-
SoftwareR: A language and environment for statistical computingR Foundation for Statistical Computing, Vienna, Austria.
-
Dissociating self-generated from passively applied head motion: Neural mechanisms in the vestibular nucleiThe Journal of Neuroscience 24:2102–2111.https://doi.org/10.1523/JNEUROSCI.3988-03.2004
-
Hearing silences: human auditory processing relies on preactivation of sound-specific brain activity patternsThe Journal of Neuroscience 33:8633–8639.https://doi.org/10.1523/JNEUROSCI.5821-12.2013
-
Gating of tactile input from the handExperimental Brain Research 79:97–102.https://doi.org/10.1007/BF00228877
-
How movement modulates hearingAnnual Review of Neuroscience 41:553–572.https://doi.org/10.1146/annurev-neuro-072116-031215
-
Short- and long-lasting consequences of novelty, deviance and surprise on brain and cognitionNeuroscience and Biobehavioral Reviews 55:268–279.https://doi.org/10.1016/j.neubiorev.2015.05.002
-
Touchant-touché: the role of self-touch in the representation of body structureConsciousness and Cognition 18:2–11.https://doi.org/10.1016/j.concog.2008.08.003
-
Optimal feedback control and the neural basis of volitional motor controlNature Reviews. Neuroscience 5:532–546.https://doi.org/10.1038/nrn1427
-
A computational neuroanatomy for motor controlExperimental Brain Research 185:359–381.https://doi.org/10.1007/s00221-008-1280-5
-
Error correction, sensory prediction, and adaptation in motor controlAnnual Review of Neuroscience 33:89–108.https://doi.org/10.1146/annurev-neuro-060909-153135
-
Evidence for sensory prediction deficits in schizophreniaThe American Journal of Psychiatry 162:2384–2386.https://doi.org/10.1176/appi.ajp.162.12.2384
-
Content-specific expectations enhance stimulus detectability by increasing perceptual sensitivityJournal of Experimental Psychology. General 144:1089–1104.https://doi.org/10.1037/xge0000109
-
A new perspective on predictive motor signalingCurrent Biology 28:R232–R243.https://doi.org/10.1016/j.cub.2018.01.033
-
Expectation in perceptual decision making: neural and computational mechanismsNature Reviews. Neuroscience 15:745–756.https://doi.org/10.1038/nrn3838
-
SoftwareJasp, version Version 0.16.3Computer Software.
-
Action enhances predicted touchPsychological Science 33:7505.https://doi.org/10.1177/09567976211017505
-
Optimal feedback control as a theory of motor coordinationNature Neuroscience 5:1226–1235.https://doi.org/10.1038/nn963
-
The JASP guidelines for conducting and reporting a Bayesian analysisPsychonomic Bulletin & Review 28:813–826.https://doi.org/10.3758/s13423-020-01798-5
-
Enhancement and suppression of tactile signals during reachingJournal of Experimental Psychology. Human Perception and Performance 43:1238–1248.https://doi.org/10.1037/xhp0000373
-
The internal structure of stopping as revealed by a sensory detection taskExperimental Brain Research 183:405–410.https://doi.org/10.1007/s00221-007-1128-4
-
Multiple paired forward and inverse models for motor controlNeural Networks 11:1317–1329.https://doi.org/10.1016/s0893-6080(98)00066-5
-
Action sharpens sensory representations of expected outcomesNature Communications 9:4288.https://doi.org/10.1038/s41467-018-06752-7
-
Action biases perceptual decisions toward expected outcomesJournal of Experimental Psychology. General 150:1225–1236.https://doi.org/10.1037/xge0000826
Article and author information
Author details
Konstantina Kilteni

Funding
Swedish Research Council (2019–01909)
- Xavier Job
- Konstantina Kilteni
HORIZON EUROPE Marie Sklodowska-Curie Actions (101059348)
- Xavier Job
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Evridiki Asimakidou and Lili Timar for their assistance during data collection. We also thank Henrik Ehrsson for his helpful comments on an earlier version of the manuscript. KK was supported by the Swedish Research Council (VR Starting Grant 2019–01909). XJ was supported by the Swedish Research Council (VR Starting Grant 2019–01909) and the European Union’s Horizon Europe research and innovation programme (Marie Skłodowska-Curie 101059348). Experimental costs were covered by the Swedish Research Council (VR Starting Grant 2019–01909).
Ethics
Human subjects: All experiments were approved by the Swedish Ethical Review Authority (registration no. 2021-03790). Participants provided written informed consent.
Copyright
© 2023, Job and Kilteni
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,171
- views
-
- 151
- downloads
-
- 26
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 26
- citations for umbrella DOI https://doi.org/10.7554/eLife.90912
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Action does not enhance but attenuates predicted touch
eLife 12:e90912.
https://doi.org/10.7554/eLife.90912




{kind=link}
{kind=link}
{kind=link}