The exon junction complex regulates the splicing of cell polarity gene dlg1 to control Wingless signaling in development
- Peking University, China
Abstract
Wingless (Wg)/Wnt signaling is conserved in all metazoan animals and plays critical roles in development. The Wg/Wnt morphogen reception is essential for signal activation, whose activity is mediated through the receptor complex and a scaffold protein Dishevelled (Dsh). We report here that the exon junction complex (EJC) activity is indispensable for Wg signaling by maintaining an appropriate level of Dsh protein for Wg ligand reception in Drosophila. Transcriptome analyses in Drosophila wing imaginal discs indicate that the EJC controls the splicing of the cell polarity gene discs large 1 (dlg1), whose coding protein directly interacts with Dsh. Genetic and biochemical experiments demonstrate that Dlg1 protein acts independently from its role in cell polarity to protect Dsh protein from lysosomal degradation. More importantly, human orthologous Dlg protein is sufficient to promote Dvl protein stabilization and Wnt signaling activity, thus revealing a conserved regulatory mechanism of Wg/Wnt signaling by Dlg and EJC.
https://doi.org/10.7554/eLife.17200.001eLife digest
Animal development involves different signaling pathways that coordinate complex behaviors of the cells, such as changes in cell number or cell shape. One such pathway involves a protein called Wingless/Wnt, which controls cell fate and growth and is also involved in tumor formation in humans. In recent decades, scientists have made a lot of progress in understanding how this signaling pathway operates. However, it is not well understood how the Wingless/Wnt signaling pathway interacts with other regulatory networks during development.
Now, Liu, Li et al. unveil a new regulatory network that controls the Wingless/Wnt pathway in fruit flies and in mammalian cells grown in the laboratory. The experiments show that an RNA binding protein family named the Exon Junction Complex positively regulates a protein called Dishevelled, which serves as a hub in the Wingless/Wnt pathway. The Exon Junction Complex keeps the amount of Dishevelled protein in check via an interaction with another protein referred to as Discs large. Further experiments indicated that Discs large binds to and protects Dishevelled from being degraded inside the cell.
Liu et al.'s findings highlight a new control mechanism for the Wingless/Wnt signaling pathway. In the future, the findings may also aid the development of new approaches to prevent or treat birth defects and cancer.
https://doi.org/10.7554/eLife.17200.002Introduction
Canonical Wingless (Wg)/Wnt signaling plays an evolutionarily conserved role in dictating cell proliferation, pattern formation, stem cell maintenance and adult tissue homeostasis. Given the importance of Wg/Wnt signaling in many cellular processes, it is not surprising that dysregulation of Wg/Wnt signaling in humans results in developmental defects as well as cancer (MacDonald et al., 2009; Clevers and Nusse, 2012). In Drosophila, Wg ligand binds to the seven-pass transmembrane receptors Frizzled/Frizzled2 (Fz/Fz2) and a co-receptor Arrow (Arr), a homolog of vertebrate LRP5/6. Formation of this trimeric complex activates a scaffold protein Dishevelled (Dsh) on the plasma membrane, leading to disruption of the Axin-mediated degradation complex and hence stabilization of Armadillo (Arm), a homolog of vertebrate β-catenin. Accumulated Arm then translocates to the nucleus to activate target gene transcription (MacDonald and He, 2012). Although core components of the Wg/Wnt signaling cascade have been identified, gaps in the understanding of this critical signaling pathway still remain to be filled. To unveil novel regulators of Wg signaling, we conducted a genome-wide RNAi screen in the developing Drosophila wing, from which a RNA binding exon junction complex (EJC) emerged as a positive regulator of Wg signaling.
The EJC is known to act in several aspects of posttranscriptional regulation, including mRNA localization, translation and degradation (Tange et al., 2004; Le Hir et al., 2016). After transcription, the pre-mRNA associated subunit eIF4AIII is loaded to nascent transcripts about 20–24 bases upstream of each exon junction, resulting in binding of Mago nashi (Mago)/Magoh and Tsunagi (Tsu)/Y14 proteins to form the pre-EJC core complex. The pre-EJC then recruits other proteins including Barentsz (Btz) to facilitate its diverse function (Shibuya et al., 2004). In vertebrates, the EJC is known to ensure translation efficiency (Nott et al., 2004) as well as to activate nonsense-mediated mRNA decay (NMD) (Gatfield et al., 2003; Chang et al., 2007). In Drosophila, however, the EJC does not contribute to NMD (Gatfield et al., 2003). It is instead required for the oskar mRNA localization to the posterior pole of the oocyte (Newmark and Boswell, 1994; Hachet and Ephrussi, 2001; Mohr et al., 2001; van Eeden et al., 2001; Palacios et al., 2004). Very recently, the pre-EJC has been shown to play an important role in alternative splicing of mRNA in Drosophila. Reduced EJC expression results in two forms of aberrant splicing. One is the exon skipping, which occurs in MAPK and transcripts that contain long introns or are located at heterochromatin (Ashton-Beaucage et al., 2010; Roignant and Treisman, 2010). The other is the intron retention on piwi transcripts (Hayashi et al., 2014; Malone et al., 2014). Furthermore, transcriptome analyses in cultured cells indicates the role of EJC in alternative splicing is also conserved in vertebrates (Wang et al., 2014).
In this study, we have utilized the developing Drosophila wing as an in vivo model system to investigate new mode of regulation of Wg signaling. We find that the pre-EJC positively regulates Wg signaling through its effect on facilitating Wg morphogen reception. Further studies reveal that the basolateral cell polarity gene discs large 1 (dlg1) is an in vivo target of the pre-EJC in Wg signaling. We show that Dlg1 acts independently from its role on cell polarity to stabilize Dsh protein, thus allowing Wg protein internalization required for signaling activation. Furthermore, we demonstrate that human Dlg2 exhibits a similar protective role on Dvl proteins to enhance Wnt signaling in cultured human cells. Taken together, our study unveils a conserved regulatory mechanism of the EJC and Dlg in Wg/Wnt signaling.
Results
The pre-EJC is required for Wg signaling in the developing Drosophila wing
The majority of the Wg/Wnt signaling components have been identified through classical forward genetic screens in Drosophila (Swarup and Verheyen, 2012; Jenny and Basler, 2014). However, these screens failed to uncover a regulatory role of RNA processing in Wg signaling, probably due to the fact that most components of RNA machineries exhibit pleiotropic effects in early development. In an in vivo RNAi screen, we found that knocking down three core components of the pre-EJC, tsu, mago and eIF4AIII, respectively, resulted in loss of marginal tissues and sensory bristles in adult Drosophila wing blade (Figure 1—figure supplement 1E–G), which resembles stereotypical phenotypes associated with reduced Wg signaling. Furthermore, loss-of-function tsuΔ18 or mago93D mutants (Roignant and Treisman, 2010) displayed similar defects in wing development (Figure 1A,B). To confirm that Wg signaling was indeed altered in pre-EJC mutants, we examined in wing imaginal discs the expression of two Wg signaling targets, senseless (sens) and Distal-less (Dll) (Seto and Bellen, 2006), which respond to graded Wg morphogen. We observed obvious loss of Sens and Dll protein production in tsu or mago somatic clones (Figure 1C–F; Figure 1—figure supplement 2A–D). However, Sens expression was not altered in somatic clones of btz (Figure 1—figure supplement 3A,B), which is a cytoplasmic component of the EJC (Palacios et al., 2004), suggesting that the role of EJC in Wg signaling is independent of its cytoplasmic function. To directly monitor transcriptional activity of Wg signaling in pre-EJC defective wing discs, two lacZ enhancer traps inserted in the genomic loci of Wg targets, frizzled3 (fz3) (Sivasankaran et al., 2000) and Dll, were used. As expected, the expression of fz3-lacZ and Dll-lacZ was decreased when tsu activity was reduced (Figure 1G,H, Figure 1—figure supplement 1H–J). Taken together, the above data indicate that the pre-EJC activity is required for Wg signaling activation in the developing fly wing.
Figure 1 with 3 supplements see all
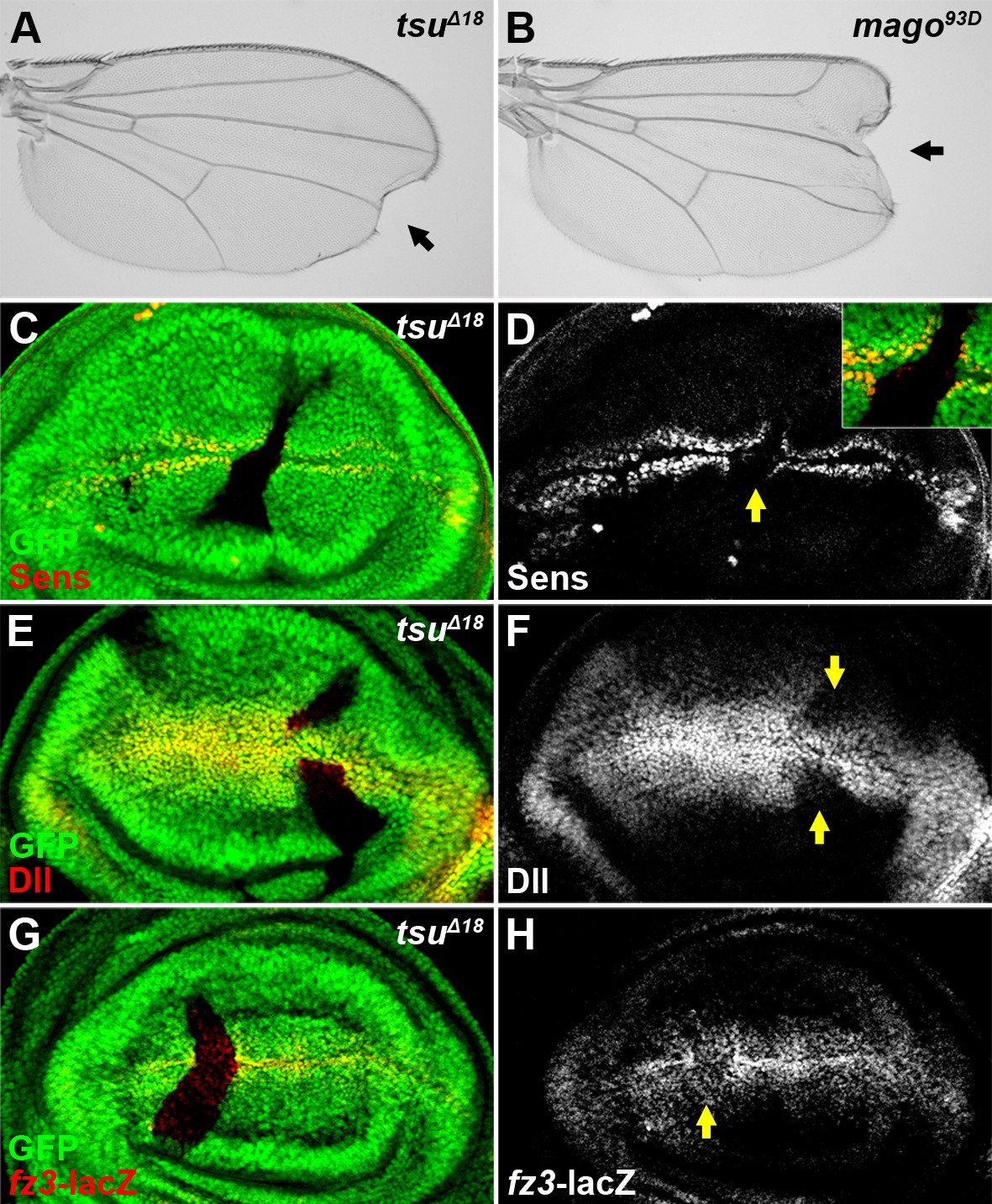
The pre-EJC positively regulates Wg signaling.
(A,B) A typical loss of Wgsignaling wing margin phenotype was observed when tsuΔ18 or mago93D mutant somatic clones were generated in adult wings. Arrows indicate serrated wing margin. (C–H) The production of Wg signaling targets Sens (C,D), Dll (E,F) and fz3-lacZ (G,H) was reduced in tsuΔ18 clones (marked by the absence of GFP and hereafter in subsequent figures). The positions of clones are indicated by arrows.
The pre-EJC regulates Wg signaling at the level of Wg protein reception
To understand how the pre-EJC regulates Wg signaling, we examined the expression of Wg signaling components in wing discs with altered EJC activity. We found that the amount of Wg protein present in tsu or mago, but not btz, mutant clones was significantly increased (Figure 2A–C; Figure 1—figure supplement 2E,F; Figure 1—figure supplement 3C,D). This finding was surprising given the observation that Wg signaling was downregulated in mutant cells. Clue to the understanding of this apparent contradiction came from the observation that Wg signaling was ectopically activated in wildtype cells immediately next to mutant clones (insets of Figures 1D and 2C). This result suggests that increased Wg protein present in mutant clones is sufficient to activate signaling in neighboring wildtype cells whilst mutant cells are incapable of receiving Wg input. Therefore, the Wg signaling defects observed in mutant cells could be caused by blockage of Wg reception at the plasma membrane. Consistent with this hypothesis, extracellular Wg protein (Strigini and Cohen, 2000) accumulated significantly in both apical and basolateral extracellular spaces (Figure 2D–F). Furthermore, we showed that the increased extracellular Wg was not due to heightened wg gene transcription nor Wg protein secretion in producing cells because neither the expression of wg-lacZ (a wg transcription reporter) nor the amount of NRT-Wg (a membrane-tethered form of Wg) was altered (Figure 2G–K; also see Figure 3—figure supplement 3B for wg transcription analyses).
Figure 2
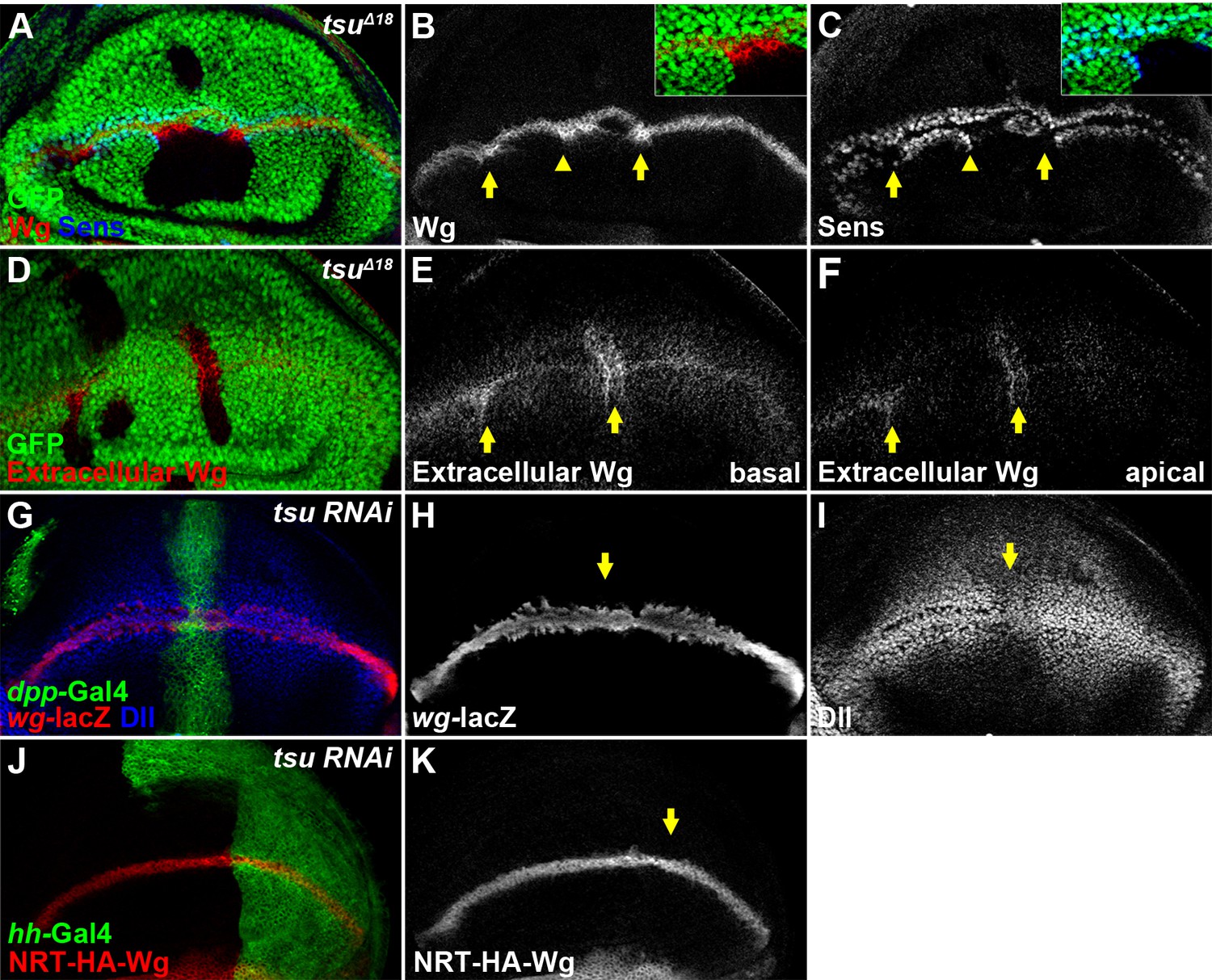
The pre-EJC is required for Wg morphogen reception.
(A–C) Wg protein stained with the conventional method (B) was accumulated in tsuΔ18 clones where the expression of Sens was reduced (arrows). The regions marked by arrowheads are shown in insets. Note that Sens was activated in cells outside the mutant clone (C). (D–F) Extracellular Wg was accumulated at the basal (E) and apical extracellular spaces of the tsuΔ18 clones (F). (G–I) The activity of wg-lacZ (H) did not change when tsu RNAi was expressed by ptc-Gal4 (marked by GFP and arrows). Note that Dll expression was reduced in tsu RNAi-expressing cells (I). (J,K) The expression of plasma membrane bound NRT-HA-Wg did not change when tsu RNAi was expressed by hh-Gal4.
Previous studies have shown that inhibiting Wg protein endocytosis by reducing either the activity of Wg receptor complex Fz2/Arr or the scaffold protein Dsh results in accumulation of Wg protein on the plasma membrane (Han et al., 2005). In tsu and mago mutant clones, we found that the expression of Dsh (Figure 3A–D), but not Fz2 or Arr (Figure 3—figure supplement 1), was obviously reduced. This result was further verified in cultured Drosophila Schneider 2 (S2) cells in which tsu or mago was knocked down by RNAi (Figure 3I). To unveil the functional importance of the EJC regulation on Dsh, overexpressed dsh was able to rescue the wing margin defects caused by tsu knockdown (Figure 3H; cf. Figure 3E), whereas fz2 or arr had little effect (Figure 3F,G). The above experiments suggest that the pre-EJC acts primarily through Dsh in wing discs to regulate Wg signaling reception.
Figure 3 with 3 supplements see all

The pre-EJC regulates Wg signaling through Dsh.
(A–D) The amount of Dsh protein was reduced in tsuΔ18 and mago69B clones in the wing disc. (E–H) Overexpressing dsh (H), but not fz2 (F) or arr (G), rescued the loss of Wg signaling wing margin phenotype caused by tsu knockdown (E). Arrows indicate sensory bristles along the wing margin. (I) The production of Myc-tagged Dsh was reduced when tsu or mago dsRNA was expressed in S2 cells. Yeast Gal80 dsRNA served as a negative control for RNAi treatment. MAPK/Rl, a known pre-EJC target served as a positive control for defective pre-EJC. β-Tubulin was used as a loading control for all experiments. (J) Real time RT-PCR revealed that the abundance of dlg1, RhoGEF2 and l(2)gl, but not dsh mRNA, was reduced when tsu dsRNA was expressed in wing discs. mapk/rl served as a positive control. α-Tubulin 84B was used to normalize the amount of cDNA template. The experiments were performed in triplicates, and data were represented as the mean+S.D. (*p<0.05; **p<0.01; Student’s t-test).
The pre-EJC regulates the splicing of cell polarity gene dlg1
The pre-EJC is known to function at the level of target mRNA splicing. However, we were not able to correlate reduced Dsh protein production observed in pre-EJC defective cells with decreased dsh mRNA expression (Figure 3J; Figure 3—figure supplement 2; Figure 3—figure supplement 3A). This inconsistency could be due to the fact that the dsh locus does not contain any intron, and in principle, may not be subjected to the EJC regulation. Thus, we reasoned that the pre-EJC must regulate Wg signaling through a Dsh-interacting protein whose mRNA expression may be controlled by the pre-EJC.
To uncover bona fide EJC targets whose encoded protein products interact with Dsh to control Wg signaling reception, we utilized whole transcriptome RNA-seq to compare mRNAs extracted from wildtype (i.e. overexpressing lacZ) and pre-EJC-defective (i.e. overexpressing tsu RNAi) wing discs, respectively. We found that the expression of 1447 mRNAs was altered by more than 25% when the pre-EJC activity was downregulated (Supplementary file 2). This list likely includes both direct as well as indirect pre-EJC targets as 629 genes were identified whose expression was increased. Among those genes whose mRNA levels were reduced we found that the expression of sens and Dll mRNA, two Wg signaling transcription targets, was decreased by more than 50% in wing discs expressing tsu RNAi. Consistent with the RT-PCR result (Figure 3—figure supplement 3), wg and dsh mRNA did not show an obvious change.
To effectively narrow down the pre-EJC targets acting in Wg signaling, we compared our candidate list with annotated information extracted from the FlyBase describing validated protein interactions with Dsh (Carmena et al., 2006; Chung et al., 2009; Kaplan and Tolwinski, 2010; Varelas et al., 2010; Johnston et al., 2013; Schertel et al., 2013; Strutt et al., 2013; Warrington et al., 2013; Garcia et al., 2014). Among 818 mRNAs whose expression was downregulated, we identified three genes, dlg1, lethal (2) giant larvae [l(2)gl] and diablo (dbo), whose protein products are known to directly interact with Dsh. qPCR and RT-PCR analyses were then performed to confirm bona fide Dsh interactors that potentially mediate the pre-EJC activity on Dsh (Figure 3J; Figure 3—figure supplement 3B). In both cases, dlg1 was the candidate consistently exhibiting reduced expression in the pre-EJC-defective wing disc cells when compared with that in wildtype cells. These results fit well with RNA-seq analyses that the amount of dlg1 mRNA was reduced by about 30% when the pre-EJC was dysfunctional. We suspected that reduced dlg1 expression may be a consequence of altered RNA splicing. Apart from previously reported exon skipping caused by dysfunctional EJC (Ashton-Beaucage et al., 2010; Roignant and Treisman, 2010), we uncovered two additional aberrant splicing events (Figure 4—figure supplement 1). The first event was exon inclusion, which retained exons that were normally efficiently spliced in specific isoforms (Figure 4—figure supplement 1E). This event likely increases the usage of splice sites. Indeed the RNA-seq analyses revealed a slightly higher usage of annotated splice sites (i.e. 1.4% higher usage for 5’ splice sites and 1.5% for 3’ splice sites) in pre-EJC depleted wing disc cells compared with those in wildtype cells (Figure 4—figure supplement 2A). The second event utilized previously unidentified splice sites to generate new introns and exons (Figure 4—figure supplement 1F–H, 2B,C; Supplementary file 3). Together, our bioinformatic analyses suggest that the pre-EJC plays a critical role in alternative splicing by preserving correct usage of splice sites to generate functional mRNA products.
The impact of the pre-EJC regulation on dlg1 splicing was further verified at the protein level as the amount of Dlg1 protein was reduced in tsu mutant wing disc clones (Figure 4A,B) as well as in S2 cells treated with dsRNA against individual components of the pre-EJC (Figure 4E). In contrast, the level of Dlg1 protein derived from a cDNA expression construct did not change (Figure 4F). Furthermore, we demonstrated that reduced dlg1 expression led to accumulation of extracellular Wg (Figure 4C,D), attenuation of Wg signaling in wing discs (Figure 4—figure supplement 3A–D), and consequently loss of wing margin and sensory bristles in the adult wing blade (Figure 4—figure supplement 3E). Significantly, overexpressed dlg1 was sufficient to rescue wing margin defects caused by dysfunctional pre-EJC (Figure 4—figure supplement 3H). It is known that additional splicing factors are required for the pre-EJC activity (Reichert et al., 2002). Inhibiting one of such factors, Rnps1, in the wing disc resulted in the same defect in Dlg1 and Dsh protein production as well as Wg signaling activation (Figure 4—figure supplement 4). Together, these experiments indicate that the pre-EJC mediated splicing activity positively regulates dlg1 to control Wg signaling in Drosophila.
Figure 4 with 6 supplements see all
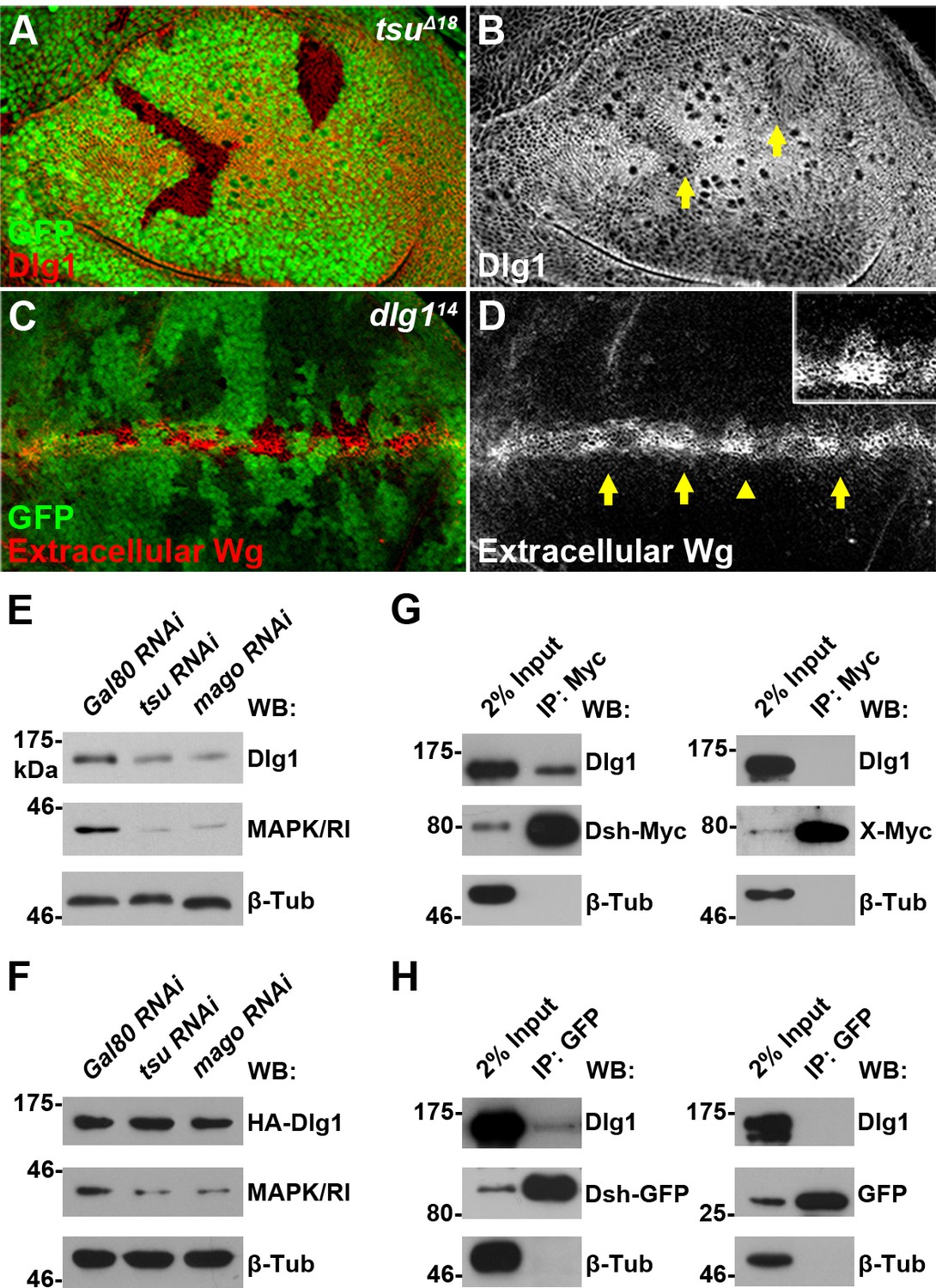
The pre-EJC regulates Dsh-interacting protein Dlg1.
(A,B) The expression of endogenous Dlg1 protein was reduced in tsuΔ18 clones (indicated by an arrow). (C,D) Extracellular Wg was accumulated in dlg114 clones. Shown in the inset is an enlarged clone with accumulated extracellular Wg. (E,F) The amount of endogenous (E) but not a cDNA-derived Dlg1 protein (F) was reduced when tsu or mago dsRNA was expressed in S2 cells. MAPK/Rl served as a positive control for dysfunctional pre-EJC activity. (G,H) Endogenous Dlg1 interacted with Myc-tagged Dsh in S2 cells (G) and GFP-tagged Dsh in wing discs expressing the dsh-gfp under the control of the dsh promoter (H). A Myc-tagged irrelevant protein X or GFP alone served as negative controls, respectively.
Dlg1 protein stabilizes Dsh in the Wg signal transduction
Dlg1 is important for the maintenance of cell polarity, acting together with L(2)gl to form a basolateral complex to counteract with the apical complex in epithelium (St Johnston and Ahringer, 2010). Therefore, it is not surprising that reduced pre-EJC activity led to altered cell polarity in wing disc epithelium (Figure 4—figure supplement 6). To investigate if the maintenance of cell polarity is required for Wg signaling, we examined the effects of L(2)gl as well as Bazooka (Baz) and Cdc42 (Figure 5—figure supplement 1), two apical complex members (Harris and Peifer, 2004; Warner and Longmore, 2009), in the Wg signal transduction. Surprisingly, loss of these polarity regulators in wing disc clones did not obviously affect Wg signaling (Figure 5—figure supplement 2), implying that the regulation of Wg signaling by Dlg1 may be independent of its function on cell polarity.
Since cell polarity does not contribute significantly to Wg signaling, we reasoned that Dlg1 may directly interact with Dsh to regulate its activity. It has been reported that Drosophila Dsh binds in vitro to the K-K-x-x-x-Ψ motif within the I3-insert of the Hook domain in Dlg1 (Garcia et al., 2014). We confirmed this interaction in S2 cells as well as in wing discs by immunoprecipitation (Figure 4G,H; Figure 4—figure supplement 6A). The relevance of such interaction was further illustrated in rescue experiments in which overexpressed dsh was able to largely rescue the wing margin defects associated with reduced dlg1 activity (Figure 4—figure supplement 3G).
Next, we investigated how Dlg1 modulates Dsh to facilitate Wg signaling. Knocking down dlg1 by RNAi in S2 cells led to reduced production of Dsh protein, whilst overexpressing dlg1 had an opposite effect (Figure 5A,B; Figure 4—figure supplement 5C). A similar result was observed in vivo when dlg1 expression was manipulated in wing discs (Figure 5C–F). The effect of Dlg1 on Dsh production may be a direct consequence of altered protein stability. Dsh protein ectopically produced in S2 cells exhibited a short half-life of around two hours (Figure 5—figure supplement 3). The turnover of Dsh protein could be controlled through the ubiquitin-proteasome or lysosome mediated degradation. We found that the degradation of Dsh protein occurred mainly in lysosome when nascent protein synthesis was blocked by cycloheximide (CHX) (Figure 5G). This is consistent with the observation that Dsh colocalized with early endosome and late endosome/lysosome markers in wing discs as well as in S2 cells (Figure 5H–M; Figure 5—figure supplement 4). Furthermore, heightened dlg1 expression counteracted with the CHX effect on Dsh degradation (Figure 5N). In contrast, Dsh degradation potentiated by tsu or dlg1 RNAi could be prevented when S2 cells were treated with a lysosomal inhibitor chloroquine (CQ) (Figure 5O,P). The above data indicate that the interaction between Dlg1 and Dsh protects Dsh protein from degradation in the lysosome. This conclusion was supported further by the observation that increased dlg1 expression correlated with reduced degree of ubiquitination modification on Dsh, a form of posttranslational modifications required for protein degradation (Figure 5Q).
Figure 5 with 4 supplements see all
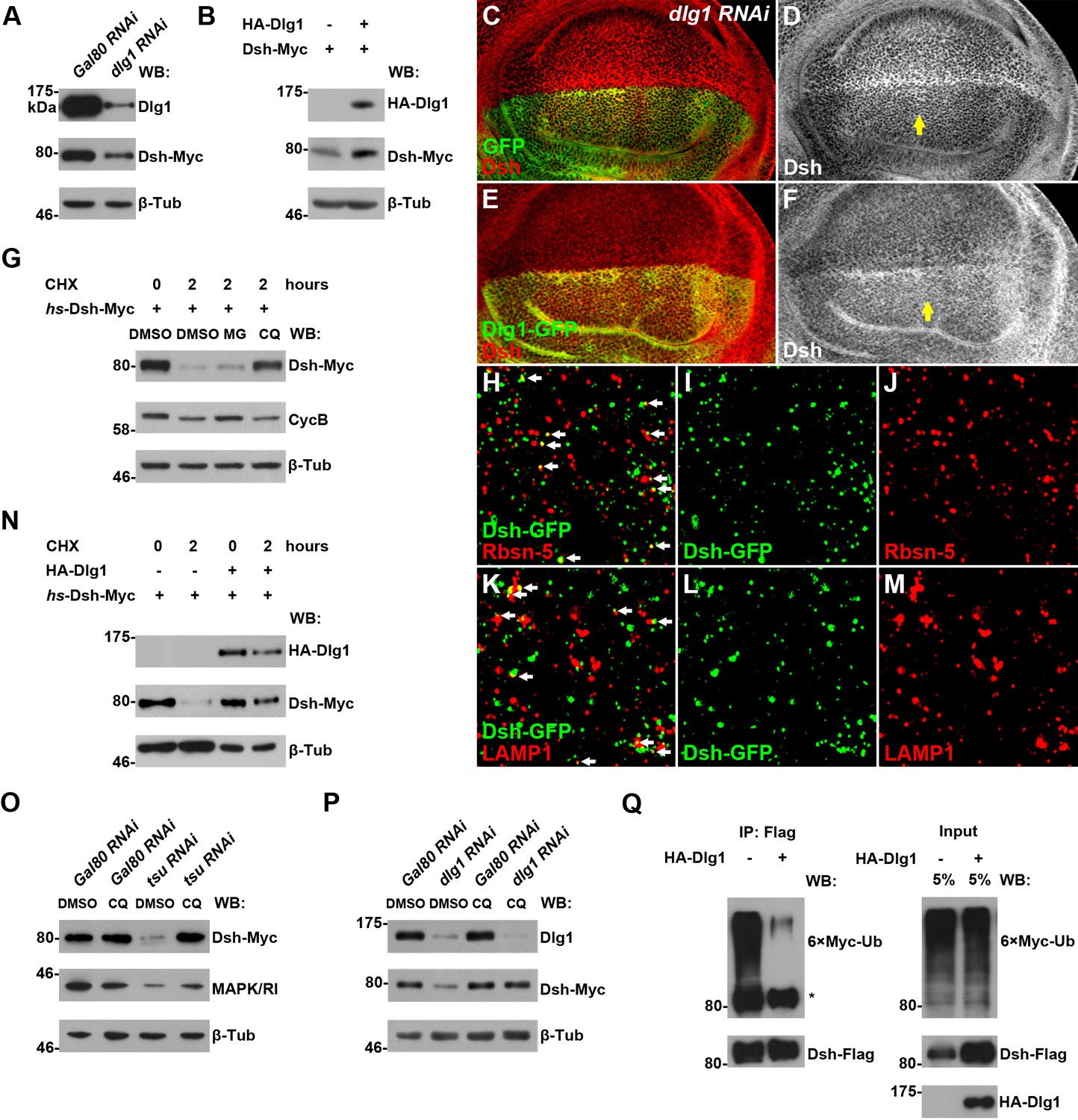
Dlg1 regulates Dsh protein turnover.
(A) Knocking down dlg1 by RNAi in S2 cells reduced the amount of Dsh protein. (B) Overexpressing dlg1 increased the abundance of Dsh protein in S2 cells. Note that Dlg1-PD form was used in all experiments unless mentioned otherwise. (C–F) The amount of Dsh was altered respectively when dlg1 RNAi (D) or dlg1-gfp (F) was expressed in wing discs by ap-Gal4. (G) CHX treatment-induced Dsh protein degradation was inhibited by lysosome inhibitor chloroquine (CQ) but not by proteasome inhibitor MG132 (MG). Cyclin B is known to be degraded in the proteasome, which served as a positive control for MG treatment (Zhang et al., 2014). DMSO served as a negative control. (H–M) Dsh protein was detected in endocytic compartments in wing discs expressing Ubpy RNAi to prevent lysosome function. Over 10% of GFP-tagged Dsh (dsh-gfp under the control of the dsh promoter) colocalized with early endosome protein Rbsn-5 [H–J; n (field of view) = 8] and late endosome/lysosome protein marker LAMP1 in wing discs [K–M; n (field of view) = 10]. (N) CHX treatment-induced Dsh degradation was inhibited when dlg1 was overexpressed in S2 cells. (O,P) Dsh degradation resulted from tsu RNAi (O) or dlg1 RNAi (P) treatment was inhibited by lysosome inhibitor CQ. (Q) Overproduced Dlg1 reduced the level of ubiquitination of Dsh. Asterisk indicated non-specific signal of Myc antibody reactivity.
The regulation of Dvl proteins by Dlg is conserved in vertebrate cells
Consistent with a role of Dlg1 on Dsh stability in Drosophila wing development, regulated protein degradation serves as a control mechanism on Dvl protein homeostasis in cultured vertebrate cells (Gao and Chen, 2010). It is thus likely that Dlg1 orthologs could act in a similar manner to control Dvl protein stability in vertebrates. Human cells encode five orthologous Dlg proteins, Dlg1-5. However, only Dlg2 contains an intact K-K-x-x-x-Ψ motif required for Dsh interaction as elucidated in Drosophila (Figure 6A). Indeed, overproduced Dlg2, but not Dlg3, was sufficient to stabilize Dvl proteins in HEK293T cells (Figure 6B,E, Figure 6—figure supplement 1A,B, Figure 6—figure supplement 2). To further illustrate the importance of the K-K-x-x-x-Ψ motif of Dlg2 in stabilizing Dvl proteins, we generated a mutant form of Dlg2 (hDlg2KKAA) where two key lysine residues of the Hook domain were mutated to alanines. As predicted, this Dlg2 mutant failed to stabilize Dvl due to its inability to interact with Dvl (Figure 6C,D). Dlg1 and Dlg2 share extensive homology within the Hook domain except that an amino acid is missing in Dlg1’s K-K-x-x-x-Ψ motif (Figure 6A). We replaced this motif in Dlg2 with amino acids found in Dlg1 (Figure 6—figure supplement 1C). The resulting Dlg2SFI-NL mutant showed reduced interaction with Dvl and no effect on Dvl protein stabilization (Figure 6—figure supplement 1D,E). To further demonstrate functional consequence of Dlg2 regulation on Dvl protein stability in Wnt signaling, we performed TOPFlash Luciferase Wnt signaling reporter assay in HEK293T cells. Overexpressed wildtype but not the KKAA mutant form of dlg2 enhanced further the Dvl-induced TOPFlash reporter activity (Figure 6F). The above experiments together indicate that the Dlg activity on Dvl protein homeostasis is conserved from flies to vertebrates.
Figure 6 with 2 supplements see all
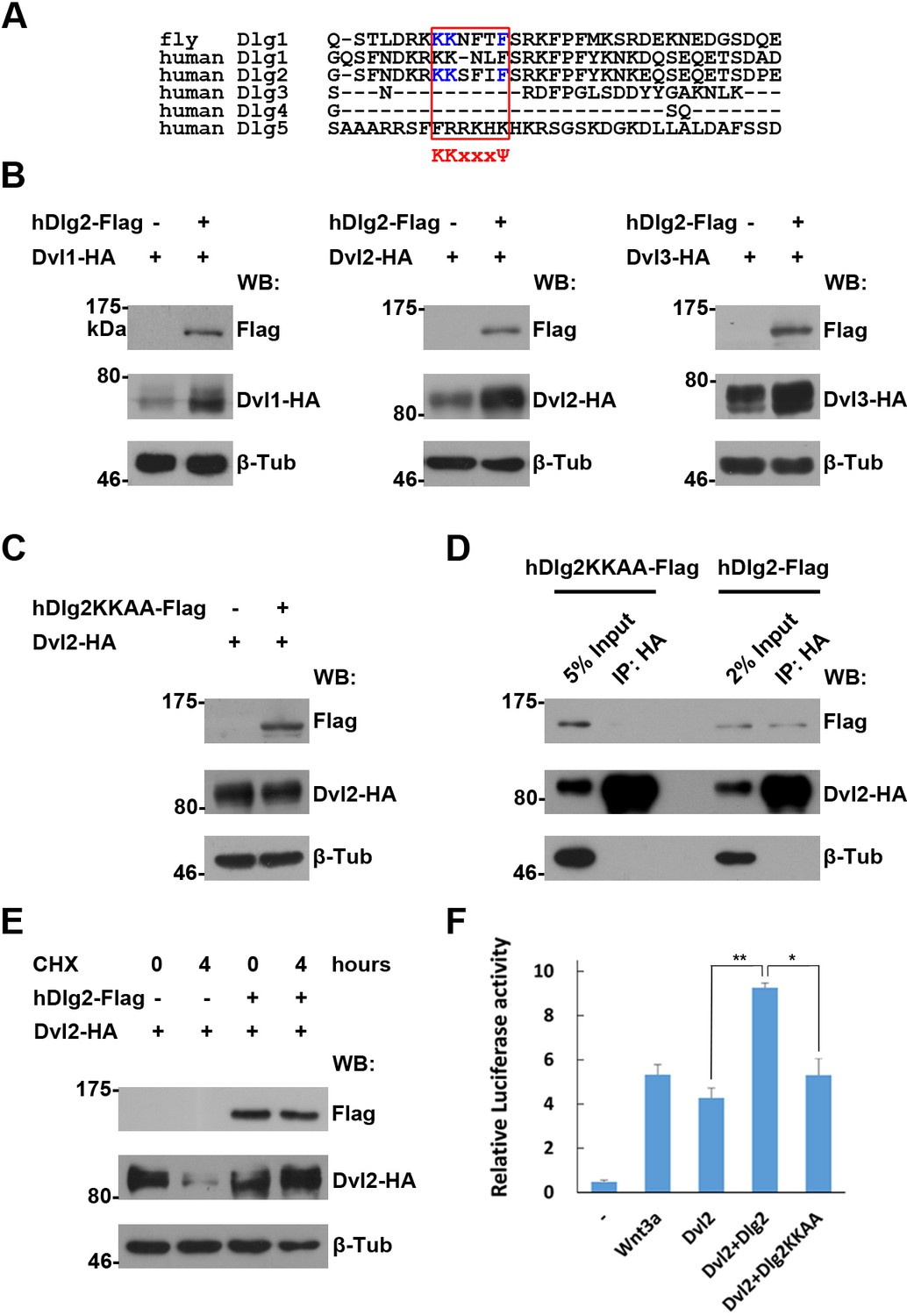
The regulation of Dvl proteins by Dlg is conserved in vertebrates.
(A) Sequence alignment of the I3-insert of Hook domains presented in Dlg proteins reveals sequence conservation between fly Dlg1 and human Dlg orthologs. The K-K-x-x-x-Ψ motif is highlighted in a red box. Amino acids essential for Dsh interaction are shown in blue. (B,C) Overproduced wildtype (B), but not the KKAA mutant form of hDlg2 (C), increased the amount of Dvl proteins in HEK293T cells. (D) Dvl protein interacted with wildtype, but not the KKAA mutant form of hDlg2. (E) CHX treatment-induced Dvl protein degradation was prevented by overproduced hDlg2. (F) Overexpressing wildtype, but not the KKAA mutant form of hDlg2, further increased Dvl2-induced TOPFlash Wnt signaling reporter activity. Ectopic Wnt3a served as a positive control. Experiments were repeated thrice, and data were represented as the mean+S.D. after normalized to Renilla activity (**p<0.01; *p<0.05; Student’s t-test).
Discussion
In summary, our study uncovers a specific role of the RNA binding protein complex EJC in the Drosophila wing morphogenesis. Our genetic and biochemical analyses demonstrate that the pre-EJC is necessary for Wg morphogen reception to activate the signal transduction. The identification of the cell polarity determinant dlg1 as one of the pre-EJC targets provides mechanistic basis for the pre-EJC regulation of the Wg signaling. Dlg1 controls the stability of the scaffold protein Dsh, which is the hub of the Wg signaling cascade (Figure 7). Importantly, this mode of regulation of Dvl by Dlg is conserved from flies to vertebrates.
Figure 7
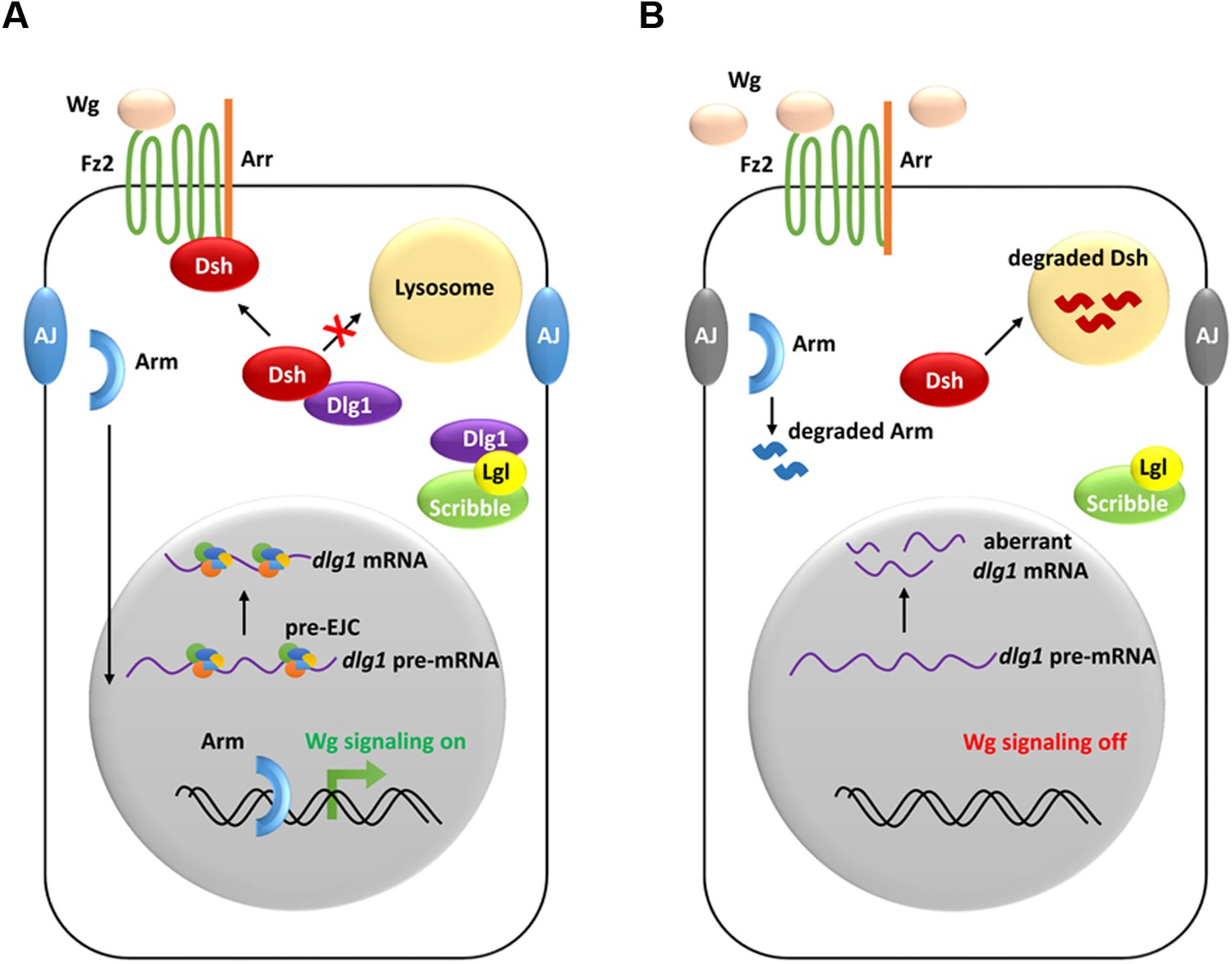
A model showing how the pre-EJC functions in the Wg signaling.
(A) In wildtype wing disc epithelial cells, the pre-EJC activity is required for precise splicing of target transcripts, including dlg1. Dlg1 protein functions independently from its role in cell polarity determination to prevent Dsh protein from degradation in the lysosome. Stabilized Dsh works together with Fz2 and Arr, two Wg co-receptors, to facilitate Wg signal reception. Consequently, through a series of cytoplasmic events, Arm protein becomes stabilized, which translocates to the nucleus to activate Wg target gene transcription. (B) When the pre-EJC activity was defective in wing disc epithelium, precise splicing of dlg1 gene is not maintained. Decreased Dlg1 production is not sufficient to stabilize Dsh, resulting in reduced Wg morphogen reception. As a result, Wg is accumulated in the extracellular space of the epithelium, which ultimately fails to protect Arm from degradation, hence reduced Wg signaling.
The EJC specifically regulates development
The EJC as well as other RNA binding protein complexes are thought to function in a pleiotropic manner. However, our data presented here together with several recent studies argue that RNA regulatory machineries can act specifically on developmental signaling for pattern formation and organogenesis. It has been increasingly recognized that the production, transport or the location of mRNA are subject to precise regulation in Wg/Wnt signaling. For example, apical localization of wg RNA is essential for signal activation in epithelial cells (Simmonds et al., 2001; Wilkie and Davis, 2001), whilst the RNA binding protein RBM47 regulates Wnt signaling in zebrafish head development (Guan et al., 2013) as well as in cancer (Vanharanta et al., 2014; Venugopal et al., 2015). The specific role of RNA machineries on cell signaling is not limited to Wg/Wnt signaling. It has been reported that RNA-binding protein Quaking specifically binds to the 3’UTR of transcription factor gli2a mRNA to modulate Hedgehog signaling in zebrafish muscle development (Lobbardi et al., 2011). RNA binding protein RBM5/6 and 10 could differentially control alternative splicing of a negative Notch regulator gene NUMB, thus antagonistically regulating the Notch signaling activity for cancer cell proliferation (Bechara et al., 2013). Therefore, generally believed pleotropic RNA regulatory machineries emerge as important regulatory means to specifically control cell signaling and related developmental processes.
The most studied function of the EJC in development is to localize oskar mRNA to the posterior pole of the oocyte for oocyte polarity establishment and germ cell formation in Drosophila (Newmark and Boswell, 1994; Hachet and Ephrussi, 2001; Mohr et al., 2001; van Eeden et al., 2001; Palacios et al., 2004). Further study suggests that the proper oskar RNA localization relies on its mRNA splicing (Hachet and Ephrussi, 2004). In light of our study of the EJC activity on dlg1 mRNA as well as the roles of EJC on mapk and piwi splicing (Ashton-Beaucage et al., 2010; Roignant and Treisman, 2010; Hayashi et al., 2014; Malone et al., 2014), we suspect that EJC might regulate oskar mRNA splicing to mediate its mRNA localization. Our RNA-seq analyses identified several hundreds of candidate mRNAs whose expression may be directly or indirectly subjected to EJC regulation. Apart from defects in Wg and MAPK signaling, however, we did not observe altered wing patterning associated with other developmental signaling systems in EJC defective flies, arguing that EJC may primarily regulate Wg and MAPK signaling in patterning the developing wing.
Regulated Dishevelled activity is required for Wg/Wnt signaling
Wg/Wnt signaling plays a fundamental role in development and tissue homeostasis in both flies and vertebrates. Its activation and maintenance rely on appropriate activity of the ternary receptor complex including Fz family proteins. In Drosophila, polarized localization of Fz and Fz2 proteins is essential for activation of non-canonical and canonical Wg signaling, respectively (Boutros et al., 2000; Wu et al., 2004). Dsh, which acts as a hub mediating both canonical and non-canonical Wg signaling, however, is found at both the apical cell boundary and in the basal side of the cytoplasm (Kaplan and Tolwinski, 2010). Thus, the polarized activity of Dsh must require distinct regulatory mechanisms at different sub-membrane compartments (Mlodzik, 2016). Our results provide the in vivo evidence suggesting that the basolateral polarity determinant Dlg1 may play a dominant role to control the Dsh abundance/activity in canonical Wg signaling.
Altered Dvl production or activity has been linked with several forms of cancer (Kafka et al., 2014). The stability of Dvl proteins can be controlled through regulated protein degradation both in vertebrates (Gao and Chen, 2010) and in Drosophila as reported in this study. In HEK293T cells, Dapper1 induces whilst Myc-interacting zinc-finger protein 1 (MIZ1) antagonizes autophagic degradation of Dvl2 in lysosome (Zhang et al., 2006; Huang et al., 2015; Ma et al., 2015). It is also reported that a tumor suppressor CYLD deubiquitinase inhibits the ubiquitination of Dvl (Tauriello et al., 2010). As Dlg1 prevents Dsh from degradation in Drosophila, it is important to investigate if Dlg1 participates in a posttranslational regulatory network of Dvl to integrate endocytosis and autophagy. Furthermore, upregulation of dvl2 and dlg2 expression has been found in various forms of cancer as shown in the COSMIC database (Forbes et al., 2015). The study of the interaction between Dlg1 and Dsh may aid the development of novel approaches to prevent or treat relevant diseases.
Complex relationship between cell polarity determination and developmental signaling
Dlg1 acts together with L(2)gl to form a basolateral complex in polarized epithelium. Dsh is known to interact with L(2)gl. On one hand, Dsh activity is required for correct localization of L(2)gl to establish apical-basal polarity in Xenopus ectoderm and Drosophila follicular epithelium (Dollar et al., 2005). On the other hand, L(2)gl can regulate Dsh to maintain planar organization of the embryonic epidermis in Drosophila (Kaplan and Tolwinski, 2010). Despite the complex interaction between L(2)gl and Dsh, not much is known about mutual regulation between Dlg1 and Dsh. A recent report suggests that Dsh binds to Dlg1 to activate Guk Holder-dependent spindle positioning in Drosophila (Garcia et al., 2014). Our results unveil another side of the relationship in which Dlg1 controls the turnover of Dsh to ensure developmental signal propagation. Apart from its apical localization at the cell boundary, Dsh is also found in the basal side of the cytoplasm (Kaplan and Tolwinski, 2010). It is likely that the interactions among Dsh, Dlg1 and L(2)gl may be dependent on their localization, and Dsh may serve as a bridge to connect cell signaling and polarity.
Developmental signaling and cell polarity intertwine to control a diverse array of cellular events. It is well known that Wg/Wnt signaling controls cell polarity in distinct manner. Non-canonical signaling acts through cytoskeletal regulators to establish planar cell polarity (Yang and Mlodzik, 2015). Canonical signaling may also directly affect apical-basal cell polarity (Karner et al., 2006). On the other hand, disruption of epithelial cell polarity has a profound impact on protein endocytosis and recycling (Barbieri et al., 2016), both of which are essential regulatory steps for signal activation and maintenance (Shivas et al., 2010). Our results add another layer of complexity by which polarity determinants could contribute to cell signaling independent of their conventional roles in polarity establishment and maintenance. Interestingly, this mode of regulation is also observed for other signaling processes. Loss of Dlg5 impairs Sonic hedgehog-induced Gli2 accumulation at the ciliary tip in mouse fibroblast cells that may not rely on cell polarity regulation (Chong et al., 2015). Similarly, L(2)gl regulates Notch signaling via endocytosis, independent of its role in cell polarity (Parsons et al., 2014). We believe that other cell polarity determinants may similarly participate in polarity-independent processes, however, the exact mechanism of how they cooperate to modulate developmental signaling awaits further investigation.
Materials and methods
Fly genetics
Request a detailed protocolThe following fly stocks were obtained from the Bloomington Drosophila Stock Center: ap-Gal4, da-Gal4, dpp-Gal4, hh-Gal4, T80-Gal4, ts-Gal80 (McGuire et al., 2003), vgBE-Gal4, baz4 FRT9-2 (#23229), cdc424 FRT19A (#9106), dlg114 FRT101 (#36283), l(2)gl27S3 FRT40A (#41561), Dll-lacZ (#10981), wg-lacZ (#1672), UAS-dsh (#9524), UAS-eIF4AIII RNAi (#32444), UAS-mago RNAi (#28931), UAS-Rnps1 RNAi (#36580), and UAS-tsu RNAi (#28955). UAS-eIF4AIII RNAi (#108580), UAS-mago RNAi (#28132), UAS-tsu RNAi (#107385) and UAS-Ubpy RNAi (#107623) were obtained from the Vienna Drosophila RNAi Center (VDRC). UAS-arr was a gift of Xinhua Lin, btz2 (Palacios and St Johnston, 2002) was a gift of Daniel St Johnston, UAS-dlg1-gfp (Zhang et al., 2007) was a gift of Bingwei Lu, dsh-gfp (Axelrod et al., 1998) was a gift of Jeffrey Axelrod, esg-flp (esg-Gal4, UAS-flp; Chen et al., 2005) and NRT-HA-wg (Alexandre et al., 2014) were gifts of Jean-Paul Vincent, UAS-fz2 (Chen and Struhl, 1999) was a gift of Gary Struhl, fz3-lacZ (Sato et al., 1999) was a gift of Roel Nusse, FRT42D mago93D and FRT42D tsuΔ18 (Roignant and Treisman, 2010) and FRT42D M(2)58F ubi-gfp were gifts of Jessica Treisman.
All fly crosses were maintained at 25°C unless noted otherwise. Detailed crossing schemes for each figure are shown in Supplementary file 1. Loss-of-function somatic clones were induced in the wing disc by Flp/FRT-mediated homologous recombination. Second instar larvae from parental crosses were heat-shocked at 37°C for half an hour. Phenotypes of the adult wing and wing discs are all fully penetrant (n>20).
Immunofluorescence staining
Request a detailed protocolFor conventional immunofluorescence staining, wing discs dissected from third instar larvae were fixed in 4% paraformaldehyde, blocked in 0.2% BSA and incubated overnight at 4°C with the following primary antibodies: rabbit anti aPKC (1:500; sc-216; Santa Cruz Biotechnology, Dallas, TX), rabbit anti-Arr (1:15000; a gift of Stephen DiNardo; Rives et al., 2006), mouse anti β-galactosidase (1:50; 40-1a; Developmental Studies Hybridoma Bank, DSHB, Iowa City, IA), rabbit anti-β-galactosidase (1:4000; Cappel, Durham, NC), mouse anti-Dlg1 (1:50; 4F3; DSHB), mouse anti-Dll (1:400; a gift of Ian Duncan; Duncan et al., 1998), rat anti-DE-Cad (1:500; DCAD2; DSHB), rat anti-Dsh (1:500; a gift from Tadashi Uemura; Shimada et al., 2001), mouse anti-Fz2 (1:20; 12A7; DSHB), mouse anti-HA (1:100; 6E2, Cell Signaling Technology, CST, Danvers, MA), rabbit anti-LAMP1 (1:500; ab30687; abcam, Cambridge, MA; Bouché et al., 2016), rabbit anti-Rbsn-5 (1:2000; a gift of Akira Nakamura; Tanaka and Nakamura, 2008), guinea pig anti-Sens (1:1000; a gift of Hugo Bellen; Nolo et al., 2000) and mouse anti-Wg (1:200; 4D4; DSHB). The wing discs were incubated with Alexa fluor-conjugated secondary antibodies (1:400; Invitrogen, Carlsbad, CA) for one hour at room temperature before mounting. Fluorescence images were acquired with a Zeiss Axio Imager Z1 microscope equipped with an ApoTome or a Leica SP8 confocal microscope. The figures were assembled in Adobe Photoshop CS5. Minor image adjustments (brightness and/or contrast) were performed in AxioVision 4.8.1 or Photoshop.
Extracellular Wg staining was performed based on a previously described protocol with minor modification (Strigini and Cohen, 2000). Briefly, third instar larvae discs were dissected in ice-cold Schneider’s Drosophila medium (Invitrogen) supplemented with 10% FBS, 100 U/ml of penicillin and 100 mg/ml of streptomycin (full medium) and then incubated on ice with mouse anti-Wg antibody (1:10; 4D4) diluted in the full medium for one hour. Larval discs were rinsed and then fixed for 20 minutes in ice-cold PBS containing 4% paraformaldehyde before proceeding for immunofluorescence staining.
To detect Dsh protein that undergoes endocytic degradation in wing discs in which dsh-gfp expression was controlled by the dsh promoter, Ubpy RNAi was used to disrupt ESCRT-0 complex that is required for delivery of internalized cargos for lysosomal degradation (Zhang et al., 2014). Similarly, S2 cells overexpressing dsh-Myc was treated with chloroquine (10 mg/ml, Sigma, St. Louis, MO) to disrupt lysosome function. Antibodies against Rbsn-5 and LAMP1 were used to label early endosomes and late endosomes/lysosomes, respectively.
Molecular biology
Request a detailed protocolMyc-tagged fly dsh was generated by cloning the dsh-Myc fragment amplified from the UAS-dsh (Bloomington #9524) transgenic strain into the NotI/XbaI site of the pUAST vector (Brand and Perrimon, 1993) or a pCaSpeR-hs vector derived from pCaSpeR. dsh-Flag construct was generated by cutting full-length fly dsh cDNA from the pUAST vector and then subcloned into the EcoRI site of a pUAST-3×Flag vector. The HA-tagged fly dlg1 was generated by fusing an HA tag at the N-terminus of the full-length dlg1-RB or dlg1-RD cDNA and then cloned into the pUAST vector. HA-tagged human dvl1, dvl2 and dvl3 were generated by fusing an HA tag at the C-terminus of the full-length human dvl1, dvl2 and dvl3 cDNA, respectively, and the resulting HA-fusions were then cloned into a pcDNA3.1 vector. Human dlg with a Flag tag at the C-terminus was generated by cloning the full-length dlg2 or dlg3 cDNA into a pCMV3×Flag vector. This vector was also used to generate the dlg2KKAA or dlg2SFI-NL mutant expression vector by site directed mutagenesis (Stratagene, La Jolla, CA). pAct-Myc-Ub plasmid was provided by Shunsuke Ishii (Dai et al., 2003). Primers used in molecular cloning are listed in Supplementary file 1.
Cell culture, transfection and RNAi treatment
Request a detailed protocolDrosophila Schneider S2 cells were cultured at 25°C in Schneider’s Drosophila full medium. HEK293T cells were grown in DMEM medium (Invitrogen) supplemented with 10% FBS, 100 µ/ml of penicillin and 100 mg/ml of streptomycin at 37°C. DNA transfection was carried out using a standard calcium phosphate protocol.
In some experiments, S2 cells or HEK293T cells were treated for up to four hours with cycloheximide (CHX; 50 µg/ml; Sigma) before harvest to inhibit nascent protein synthesis. MG132 (50 µM; Sigma) was used to inhibit the proteasome activity, while chloroquine (10 mg/ml) was used to inhibit lysosome function (Zhang et al., 2014). S2 cells transfected with hs-dsh and indicated vectors were heat shock for half an hour at 37°C after transfection for 48 hours. Then the cells were recovered at 25°C for one hour followed by drug treatment.
dsRNA was generated with the MEGAscript high yield transcription kit (Ambion, Austin, TX) according to the manufacturer’s instruction. DNA template targeting tsu (encoding amino acids 45–165), mago (encoding amino acids 31–140), dlg1 (encoding amino acids 800–917) and full length yeast Gal80 was generated by PCR and used for dsRNA synthesis. dsRNA targeting yeast Gal80 coding sequence was used as a negative control (Su et al., 2011). For RNAi knockdown in S2 cells, dsRNA transfection was carried out using a standard calcium phosphate protocol. Primers used to generate dsRNAs are listed in Supplementary file 1.
TOPFlash Wnt signaling luciferase reporter assay
Request a detailed protocolHEK293T cells grown in 24 well plates were transfected with the TOPFlash luciferase reporter (a gift of Yeguang Chen; Zhang et al., 2006) and indicated vectors for two days before harvest. The pRL-TK Renilla reporter was co-transfected to normalize transfection efficiency. Luciferase activity was measured following the Dual-Glo luciferase assay protocol (Promega, Madison, WI).
Immunoblotting, immunoprecipitation and ubiquitination assays
Request a detailed protocolS2, HEK293T cells and 3rdinstar larval wing imaginal discs were lysed in NP-40 buffer (1% NP-40, 150 mM NaCl and 50 mM Tris-Cl, pH 8) supplemented with protease inhibitor cocktail (Roche, Germany). The concentration of protein cell lysate was quantified using a BCA protein assay kit (Thermo, Waltham, MA). Immunoblotting was carried out using standard protocols. The following antibodies were used for immunoblotting: mouse anti-β-Tubulin (1:10000; E7, DSHB), mouse anti-Cyclin B (1:50; F2F4; DSHB), mouse anti-Dlg1 (1:2000; 4F3; DSHB), rabbit anti-Flag tag (1:2000; D6W5B; CST), rabbit anti-HA tag (1:1000; Y-11; Santa Cruz), mouse anti-HA tag (1:2000; 6E2; CST), rabbit anti-MAPK (1:1000, 137F5, CST) and mouse anti-Myc tag (1:2000; 9B11; CST).
Immunoprecipitation was performed using agarose anti-HA (Vector Labs, Burlingame, CA), agarose anti-Myc (Vector Labs), agarose anti-GFP (Vector Labs) or Flag M2 affinity gel (Sigma) according to manufacturers’ instructions. Immunoblots presented in all figures are representatives of at least three independent experiments.
Ubiquitination assays were carried out with hot lysis-extracted protein lysates based on the protocol described previously (Row et al., 2006; Zhang et al., 2014). Briefly, S2 cells transfected with dsh-Flag, HA-dlg and 6×Myc-Ub were hot-lysed in denaturing buffer (1% SDS, 50 mM Tris, pH 7.5, 0.5 mM EDTA) by boiling for five minutes. Lysates were then diluted 10-fold with NP-40 lysis buffer and subject to immunoprecipitation with anti-Flag M2 affinity gel (Sigma).
RNA isolation, quantitative real-time PCR and RT-PCR
Request a detailed protocolTotal RNA of pooled third instar larvae or imaginal wing discs was extracted using TRIzol reagent (Invitrogen). Residual genomic DNA was removed by RNase-free DNase (New England Biolabs, NEB, Ipswich, MA). First strand cDNA was synthesized using oligo-dT primers and SuperScript III reverse transcriptase (Invitrogen). Quantitative real-time PCR was performed using SYBR Green PCR master mix (Applied Biosystems, Waltham, MA) on a 7500 real time PCR system (Applied Biosystems). Using α-Tubulin 84B as an internal control, relative fold changes of transcripts were calculated using comparative CT (2–ΔΔCT) method. Three independent samples were prepared and run in triplicates. RT-PCR was performed to compare with the quantitative real-time PCR results. Primers used in quantitative real-time PCR and RT-PCR are listed in Supplementary file 1.
Whole transcriptome sequencing and analyses
Request a detailed protocolTotal RNA from pooled third instar larval wing imaginal discs (1000 pairs per sample preparation) expressing UAS-lacZ or UAS-tsu RNAi (VDRC#107385) driven by T80-Gal4 was extracted in duplicates using TRIzol. Poly(A)+ mRNAs were enriched using Dynabeads oligo (dT) beads (NEB). RT reactions and purification of cDNA templates were performed following the RNA-seq sample preparation protocol from Illumina. Each cDNA sample was sequenced on an IlluminaHiseq 2500 system.
Whole transcriptome reads were aligned using the TopHat (v2.0.13) (Trapnell et al., 2012) with the Ensembl Drosophila_melanogaster.BDGP5.78.gtf as a reference (http://www.ensembl.org/index.html). In total, 22,563,753 and 24,660,199 of 125 bp reads pairs for duplicated tsu samples and 22,610,210 and 28,651,178 of 125 bp reads pairs for lacZ samples were sequenced, respectively. The transcription analysis was performed using Cufflinks (v2.2.1) (Trapnell et al., 2012). DEXseq was used to plot the transcripts to each gene (Anders et al., 2012). RPKM method (reads per kilobase of transcript per million mapped sequence reads; Mortazavi et al., 2008) was used for normalizing gene counts. We calculated the ratio of RPKM between the tsu and lacZ samples. The R density plot package (R Core Team, 2015, R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org//) was used to generate the distribution plot shown in Figure 4—figure supplement 2C. The Seq data was deposited to NCBI website: http://www.ensembl.org/index.html http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token=ujyhqauybbujhun&acc=GSE81220.
Data availability
-
The exon junction complex regulates the splicing of cell polarity gene dlg1 to control Wingless signaling in developmentPublicly available at the NCBI Gene Expression Omnibus (accession no: GSE81220).
References
-
Detecting differential usage of exons from RNA-seq dataGenome Research 22:2008–2017.https://doi.org/10.1101/gr.133744.111
-
Endocytic control of signaling at the plasma membraneCurrent Opinion in Cell Biology 39:21–27.https://doi.org/10.1016/j.ceb.2016.01.012
-
Targeted gene expression as a means of altering cell fates and generating dominant phenotypesDevelopment 118:401–415.
-
The nonsense-mediated decay RNA surveillance pathwayAnnual Review of Biochemistry 76:51–74.https://doi.org/10.1146/annurev.biochem.76.050106.093909
-
Wingless transduction by the Frizzled and Frizzled2 proteins of DrosophilaDevelopment 126:5441–5452.
-
Allocation and specification of the genital disc precursor cells in DrosophilaDevelopmental Biology 281:270–285.https://doi.org/10.1016/j.ydbio.2005.02.032
-
Bifurcating action of Smoothened in Hedgehog signaling is mediated by Dlg5Genes & Development 29:262–276.https://doi.org/10.1101/gad.252676.114
-
COSMIC: exploring the world's knowledge of somatic mutations in human cancerNucleic Acids Research 43:D805–811.https://doi.org/10.1093/nar/gku1075
-
Dishevelled: The hub of Wnt signalingCellular Signalling 22:717–727.https://doi.org/10.1016/j.cellsig.2009.11.021
-
rbm47, a novel RNA binding protein, regulates zebrafish head developmentDevelopmental Dynamics 242:1395–1404.https://doi.org/10.1002/dvdy.24039
-
The cellular story of dishevelledsCroatian Medical Journal 55:459–467.https://doi.org/10.3325/cmj.2014.55.459
-
Spatially defined Dsh-Lgl interaction contributes to directional tissue morphogenesisJournal of Cell Science 123:3157–3165.https://doi.org/10.1242/jcs.069898
-
Apical-basal polarity, Wnt signaling and vertebrate organogenesisSeminars in Cell & Developmental Biology 17:214–222.https://doi.org/10.1016/j.semcdb.2006.05.007
-
The exon junction complex as a node of post-transcriptional networksNature Reviews Molecular Cell Biology 17:41–54.https://doi.org/10.1038/nrm.2015.7
-
The Wnt signaling antagonist Dapper1 accelerates Dishevelled2 degradation via promoting Its ubiquitination and aggregate-induced autophagyJournal of Biological Chemistry 290:12346–12354.https://doi.org/10.1074/jbc.M115.654590
-
Frizzled and LRP5/6 Receptors for Wnt/β-Catenin SignalingCold Spring Harbor Perspectives in Biology 4:a007880.https://doi.org/10.1101/cshperspect.a007880
-
Wnt/β-catenin signaling: components, mechanisms, and diseasesDevelopmental Cell 17:9–26.https://doi.org/10.1016/j.devcel.2009.06.016
-
The Dishevelled protein family: still rather a mystery after over 20 years of molecular studiesCurrent Topics in Developmental Biology 117:75–91.https://doi.org/10.1016/bs.ctdb.2015.11.027
-
Mapping and quantifying mammalian transcriptomes by RNA-SeqNature Methods 5:621–628.https://doi.org/10.1038/nmeth.1226
-
The mago nashi locus encodes an essential product required for germ plasm assembly in DrosophilaDevelopment 120:1303–1313.
-
SoftwareR: A language and environment for statistical computingR Foundation for Statistical Computing, Vienna, Austria.
-
The ubiquitin isopeptidase UBPY regulates endosomal ubiquitin dynamics and is essential for receptor down-regulationJournal of Biological Chemistry 281:12618–12624.https://doi.org/10.1074/jbc.M512615200
-
Dfrizzled-3, a new Drosophila Wnt receptor, acting as an attenuator of Wingless signaling in wingless hypomorphic mutantsDevelopment 126:4421–4430.
-
Internalization is required for proper Wingless signaling in Drosophila melanogasterJournal of Cell Biology 173:95–106.https://doi.org/10.1083/jcb.200510123
-
eIF4AIII binds spliced mRNA in the exon junction complex and is essential for nonsense-mediated decayNature Structural & Molecular Biology 11:346–351.https://doi.org/10.1038/nsmb750
-
Polarity and endocytosis: reciprocal regulationTrends in Cell Biology 20:445–452.https://doi.org/10.1016/j.tcb.2010.04.003
-
The Wingless target gene Dfz3 encodes a new member of the Drosophila Frizzled familyMechanisms of Development 91:427–431.https://doi.org/10.1016/S0925-4773(99)00313-5
-
Wingless gradient formation in the Drosophila wingCurrent Biology 10:293–300.https://doi.org/10.1016/S0960-9822(00)00378-X
-
Wnt/Wingless signaling in DrosophilaCold Spring Harbor Perspectives in Biology 4:007930.https://doi.org/10.1101/cshperspect.a007930
-
The ever-increasing complexities of the exon junction complexCurrent Opinion in Cell Biology 16:279–284.https://doi.org/10.1016/j.ceb.2004.03.012
-
The Hippo pathway regulates Wnt/β-catenin signalingDevelopmental Cell 18:579–591.https://doi.org/10.1016/j.devcel.2010.03.007
-
Distinct functions for Rho1 in maintaining adherens junctions and apical tension in remodeling epitheliaJournal of Cell Biology 185:1111–1125.https://doi.org/10.1083/jcb.200901029
-
Wnt-Frizzled/planar cell polarity signaling: cellular orientation by facing the wind (Wnt)Annual Review of Cell and Developmental Biology 31:623–646.https://doi.org/10.1146/annurev-cellbio-100814-125315
-
Dapper 1 antagonizes Wnt signaling by promoting Dishevelled degradationJournal of Biological Chemistry 281:8607–8612.https://doi.org/10.1074/jbc.M600274200
Article and author information
Author details
Funding
State Key Laboratory of Membrane Biology of the People's Republic of China
- Alan Jian Zhu
Peking-Tsinghua Center for Life Sciences
- Cheng Li
- Alan Jian Zhu
Ministry of Science and Technology of the People's Republic of China (2014CB942804)
- Alan Jian Zhu
National Science Foundation of the People's Republic of China (31371410)
- Alan Jian Zhu
National Science Foundation of the People's Republic of China (31401241)
- Min Liu
China Postdoctoral Science Foundation (Postdoctoral Fellowship, 2014M550556)
- Min Liu
Peking-Tsinghua Center for Life Sciences (Postdoctoral Fellowship)
- Min Liu
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank everyone who shared with us reagents, and the Bloomington Drosophila Stock Center, the Vienna Drosophila RNAi center (VDRC) and the Developmental Studies Hybridoma Bank (DSHB) for fly stocks and antibodies.
Copyright
© 2016, Liu et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,400
- views
-
- 547
- downloads
-
- 12
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 12
- citations for umbrella DOI https://doi.org/10.7554/eLife.17200
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The exon junction complex regulates the splicing of cell polarity gene dlg1 to control Wingless signaling in development
eLife 5:e17200.
https://doi.org/10.7554/eLife.17200




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}