Structure–function analysis of Lactiplantibacillus plantarum DltE reveals D-alanylated lipoteichoic acids as direct cues supporting Drosophila juvenile growth
- Molecular Microbiology and Structural Biochemistry, CNRS UMR 5086, Université Claude Bernard Lyon 1, France
- Institut de Génomique Fonctionnelle de Lyon, Ecole Normale Supérieure de Lyon, CNRS UMR 5242, Université Claude Bernard Lyon 1, France
- Université Paris-Saclay, INRAE, AgroParisTech, Micalis Institute, France
- Protein Science Facility, CNRS UAR3444, INSERM US8, Université Claude Bernard Lyon 1, Ecole Normale Supérieur de Lyon, France
- Institute for Glyco-core Research (iGCORE), Gifu University, Japan
- Univ. Lille, CNRS, UMR 8576 - UGSF - Unité de Glycobiologie Structurale et Fonctionnelle, France
Abstract
Metazoans establish mutually beneficial interactions with their resident microorganisms. However, our understanding of the microbial cues contributing to host physiology remains elusive. Previously, we identified a bacterial machinery encoded by the dlt operon involved in Drosophila melanogaster’s juvenile growth promotion by Lactiplantibacillus plantarum. Here, using crystallography combined with biochemical and cellular approaches, we investigate the physiological role of an uncharacterized protein (DltE) encoded by this operon. We show that lipoteichoic acids (LTAs) but not wall teichoic acids are D-alanylated in Lactiplantibacillus plantarumNC8 cell envelope and demonstrate that DltE is a D-Ala carboxyesterase removing D-Ala from LTA. Using the mutualistic association of L. plantarumNC8 and Drosophila melanogaster as a symbiosis model, we establish that D-alanylated LTAs (D-Ala-LTAs) are direct cues supporting intestinal peptidase expression and juvenile growth in Drosophila. Our results pave the way to probing the contribution of D-Ala-LTAs to host physiology in other symbiotic models.
Editor's evaluation
This is an important study on the role of a bacterial cell wall component, D-alanylated lipoteichoic acid, as a cue in Drosophila melanogaster-microbiome interactions. Overall, the evidence presented to support the conclusions is compelling. The approach combines crystallography with biochemical and cellular assays that take advantage of both fly and bacterial mutants to demonstrate a physiological role in juvenile growth promotion. The work will be of broad interest to those studying host-microbe interactions, particularly aspects related to immunology and metabolism, mediated by the microbiome.
https://doi.org/10.7554/eLife.84669.sa0Introduction
Metazoans establish mutually beneficial interactions with their resident microorganisms (McFall-Ngai et al., 2013). These interactions contribute to different aspects of host physiology, including juvenile growth, a postnatal developmental process marked by rapid body-size increase and organ maturation (Schwarzer et al., 2018). Juvenile growth results from the integration of environmental cues with the organism’s intrinsic genetic potential, driven by energy and nutritional demands. Harsh environmental conditions, notably nutrient deprivation, result in linear and ponderal growth failure (i.e. stunting) (Nabwera et al., 2022), as well as alteration in gut microbiota maturation (Subramanian et al., 2014), which can be mitigated by microbial interventions and/or microbiota-directed nutritional interventions (Barratt et al., 2022). Despite some recent advances, the understanding of how gut microbes buffer the deleterious effect of undernutrition and contribute to healthy juvenile growth remains elusive.
Drosophila melanogaster (referred here as Drosophila) is a valuable experimental model to study the physiological consequences and underlying mechanisms of host–commensal bacteria interactions (Douglas, 2018; Grenier and Leulier, 2020). Bacterial strains associated with Drosophila influence multiple physiological processes, including juvenile growth, and on several occasions, the underlying symbiotic cues, that is, bacterial molecules directly impacting host functionalities, have been identified. For instance, amino acids produced by symbiotic bacteria can inhibit the production of the neuropeptide CNMamide in the gut, which shape food foraging behavior by repressing preference for amino acids (Kim et al., 2021). Bacterial metabolite such as acetate produced by strains of Acetobacter pomorum is necessary to support juvenile growth (Shin et al., 2011) by altering the epigenome of enteroendocrine cells and stimulating the secretion of the intestinal hormone Tachykinin (Jugder et al., 2021; Kamareddine et al., 2018). In the context of microbe-mediated Drosophila juvenile growth promotion (Storelli et al., 2011), peptidoglycan (PG) fragments from Lactiplantibacillus plantatum (Lp) cell walls are directly sensed by peptidoglycan recognition receptors (PGRPs) in Drosophila enterocytes. This recognition signal, via the IMD/NF-kappaB pathway, promotes the production of intestinal peptidases, which helps juveniles optimizing the assimilation of dietary proteins to support their systemic growth (Erkosar et al., 2015).
Recently, in an effort to further characterize the bacterial machinery involved in Lp-mediated juvenile growth promotion, we identified through forward genetic screening the pbpX2-dltXABCD operon as an important determinant of Lp-induced Drosophila larval growth (Matos et al., 2017). The first gene of the operon, pbpX2 (here renamed dltE for D-Ala-LTA Esterase, see below), is uncharacterized and annotated as a serine-type D-Ala-D-Ala-carboxypeptidase putatively involved in PG maturation cleaving the terminal D-alanine (D-Ala) residue of the peptide stem in newly made muropeptides. Remarkably, Lp PG precursors do not contain a terminal D-Ala in peptide stems but a terminal D-lactate (D-Lac) (Ferain et al., 1996) raising the question of the biochemical activity of DltE. On the other hand, the remaining dltABCD encode a multi-protein machinery responsible for the D-alanylation of teichoic acids (TA) in diverse Gram-positive bacteria (Perego et al., 1995) and in Lp (Nikolopoulos et al., 2022). TAs are anionic polymers localized within the Gram-positive bacteria cell wall, representing up to 50% of the cell envelope dry weight and present in two forms: wall teichoic acids (WTAs), which are covalently bound to PG, and lipoteichoic acids (LTAs), which are anchored to the cytoplasmic membrane (Rohde, 2019). Study of an isogenic mutant of Lp NC8 (LpNC8) strain, deleted for the entire dlt operon, revealed that this operon is indeed essential to D-Ala esterification to the cell envelope (most likely on TAs) and that purified D-alanylated cell envelopes triggered intestinal peptidase expression and support Drosophila juvenile growth (Matos et al., 2017). These results suggested that TA modifications are important cues shaping commensal-host molecular dialog. However, which type of TAs and whether their modification is directly involved remains unaddressed and an indirect influence of the D-alanylation process on PG maturation and PG sensing by the host could not be excluded.
To address these standing questions, we investigate here the structure, the biochemical activity, and the physiological role of DltE. We also study the impact of DltE on TA structure and modifications in Lp cell envelope and test the relative contribution of purified PG, WTA, and D-alanylated-LTA (D-Ala-LTA) from Lp cell envelope to support Drosophila growth. Our results establish that DltE is not a carboxypeptidase modifying Lp PG but rather a D-Ala esterase acting upon D-Ala-LTA. After characterizing the chemical structure of LTAs and WTAs, we show that only LTAs but not WTAs are D-alanylated in LpNC8 cell envelopes and we demonstrate that D-Ala-LTAs, in addition to PG, are direct cues supporting intestinal peptidase expression and juvenile growth in Drosophila.
Results
Structure of the extracellular domain of DltE
DltE is annotated as a putative serine type D-Ala-D-Ala carboxypeptidase. However, sequence comparison with canonical members of this protein family revealed low sequence identity (Figure 1—figure supplement 1). Additionally, while the catalytic S-X-X-K motif (Motif 1) is conserved in DltE, the second (Y-X-S) and third (K/H-T/S-G) motifs that complete the active site are altered with only the catalytic Tyr and the Gly residues being conserved, respectively. This suggests a modified active site environment with potentially different catalytic and/or substrate binding mechanisms. Supporting this, we showed that the extracellular catalytic domain of DltE (DltEextra 34–397) is not able to bind penicillin although this property is a hallmark of serine type D-alanyl-D-alanine carboxypeptidases that belongs to the PBP (penicillin binding proteins) protein family (Figure 1—figure supplement 2).
These features prompted us to determine the structure of the extracellular domain of DltE (DltEextra) by X-ray crystallography (Supplementary file 1). The domain consists of two subdomains, an α-β sandwich (residues 76–126 and 246–397) that form a five strand (β1–β5) antiparallel β-sheet flanked by 5 α-helices (α2, α9–α12) and an α-helix-rich region (residues 127–245) folding in 6 α-helices (α3-α8) (Figure 1a and Figure 1—figure supplement 1). DltE is structurally homologous to the D-Ala-D-Ala carboxypeptidase R61 (DDCP) from Streptomyces (Figure 1b and c; Kelly et al., 1986; McDonough et al., 2002) that exhibits the classical β-lactamase fold of PBPs (Figure 1b). The positions of the active site S-X-X-K motif (128SIQK131), containing the catalytic Ser128 and Lys131 residues, and the catalytic Tyr213 from the second motif are conserved (Figure 1a and b and Figure 2). These last two residues are expected to function as a general base during the acylation step and function as a relay mechanism for the transfer of a proton from the incoming substrate to the departing catalytic serine. In contrast, several striking features distinguish DltE from DDCP and more generally from PBPs and DD-carboxypeptidases. The β3-strand that usually bears the third motif and defines one side of the catalytic cavity is not conserved in terms of length and position (McDonough et al., 2002; Figure 1c). Second, DltE contains an extra α-helix (α1) at the N-terminus of the catalytic domain (Figure 1c), as well as three major structural differences affecting the regions lining the sides of the active site of DDCP and responsible for its cavity-like shape (Figure 1c, e and g): (1) a 15-amino acid long loop connecting α4 and α5 in DltE (residues 175–190 named Loop I on Figure 1c and d and Figure 1—figure supplement 1) that replaces a longer region of 20 residues that is folded in α-helices in DDCP; (2) the mostly unstructured segment ranging from residues 328–346 in DltE and located between α11 and β3 (Loop II on Figure 1c and d and Figure 1—figure supplement 1) is 10 residues longer in DDCP in which it forms an antiparallel β-sheet of three small strands that follows the main 5-stranded β-sheet; and (3) the loop between α8 and α9 (residues 257–283 in DltE) connecting the two subdomains is slightly shorter in DltE and adopts a different position (Figure 1c). This last difference is of particular interest because this loop was described as the Ω-like loop in β-lactamases and PBP proteins (Fetrow, 1995; Yi et al., 2016) and shown to play a key role in the maintenance of the active site topology and in the enzymatic activity (Figure 1e and Figure 1—figure supplement 1). The global architecture of the active site cavity of DltE is therefore reshaped and different from that of DDCP enzymes, adopting a deeper cleft topology largely opened on both sides of the protein (Figure 1d–g). Together with its inability to bind penicillin, such structural features strongly suggest that DltE possesses an alternative substrate recognition mode and/or a different substrate specificity than PBPs.
Figure 1 with 2 supplements see all
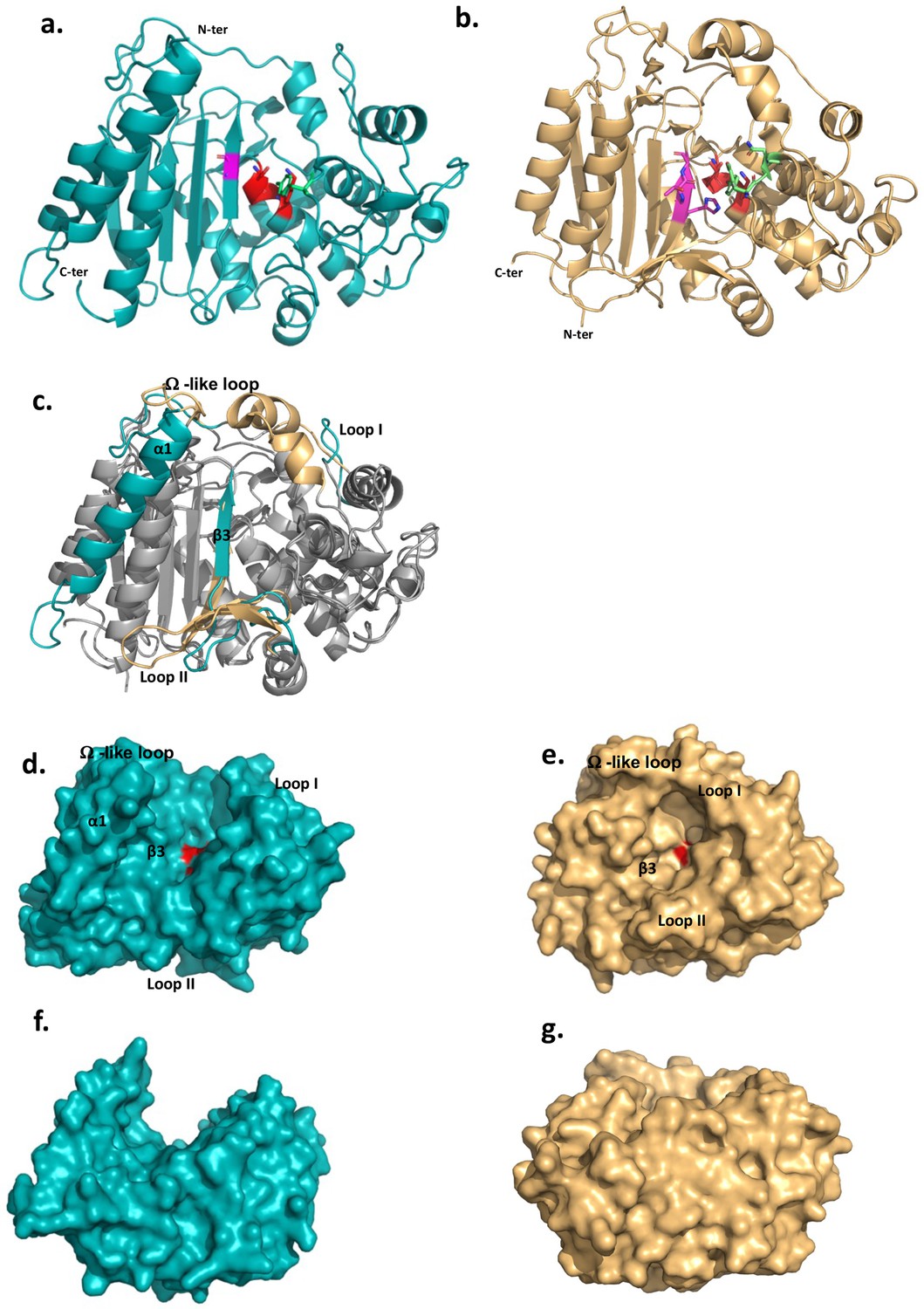
The 3D structures of L. plantarum DltEextra and its structural comparison with the Streptomyces R61 D-Ala-D-Ala carboxypeptidase structure.
(a) Cartoon representations of the 3D X-ray structure of L. plantarum DltEextra and (b) of the canonical D-Ala-D-Ala carboxypeptidase from Streptomyces sp. R61 (DDCP) (PDB ID 1HVB/1IKG). The N- and C-termini are indicated. The canonical DDCP conserved motifs 1 (SXXK, with S the catalytic Ser), 2 (YXN), and 3 ((K/H)(S/T)G) are highlighted in red, green, and pink, respectively. The first motif is strictly conserved in DltE (128SIQK131) and located as for DDCP at the beginning of the α-helix-rich region (α3 helix in DltE, α2 in DDCP) at the interface with the β-sheet. Only the Tyr and Gly residues are conserved in the motifs 2 and 3, respectively. The position of the catalytic Tyr213 from the second motif is also conserved and found in the loop connecting α5 and α6, close to the catalytic dyad. (c) Superimposition between DltEextra and DDCP (PDB ID 1HVB/1IKG) 3D structures. The common structural cores that exhibit a classical β-lactamase fold or penicillin binding (PB) fold are colored in gray. They are well superimposed with rms deviation calculated at 2.2 Å on around 300 residues. The main structural differences are highlighted in teal for DltE and wheat for DDCP and indicated on the figure: the N-terminal α1 helix of DltE, the β3-strand that contains the motif 3, the Loop I, the Loop II, and the Ω-like loop. (d, f) and (e, g) Surface representation of DltEextra and DDCP. (d, e) and (f, g) are shown in the same orientation. (f) and (g) are rotated by 90° along a horizontal axis compared to (d) and (e), respectively. The catalytic Ser, colored in red, is buried at the bottom of the catalytic cavity in DDCP and lying in the middle of a large cleft in DltE. The structural elements that define the active site architecture and differ between DltE and DDCP are indicated.
-
Figure 1—source data 1
DltE 3D structure.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data1-v2.xlsx
-
Figure 1—source data 2
Raw SDS-PAGE analysis of the purity of DltEextra.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data2-v2.pdf
-
Figure 1—source data 3
Labeled SDS-PAGE analysis of the purity of DltEextra.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data3-v2.pdf
-
Figure 1—source data 4
Size-exclusion chromatography (SEC) of the Ni-affinity purified DltEextra on a Superdex 200 10/300 GL.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data4-v2.xlsx
-
Figure 1—source data 5
Microscale thermophoresis (MST) normalized dose–response data for the binding interaction between penicillin and S. pneumoniae PBP2b.
The experiment was made in triplicate.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data5-v2.xlsx
-
Figure 1—source data 6
Microscale thermophoresis (MST) normalized dose–response data for the binding interaction between penicillin and DltEextra.
The experiment was made in triplicate.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig1-data6-v2.xlsx
Figure 2
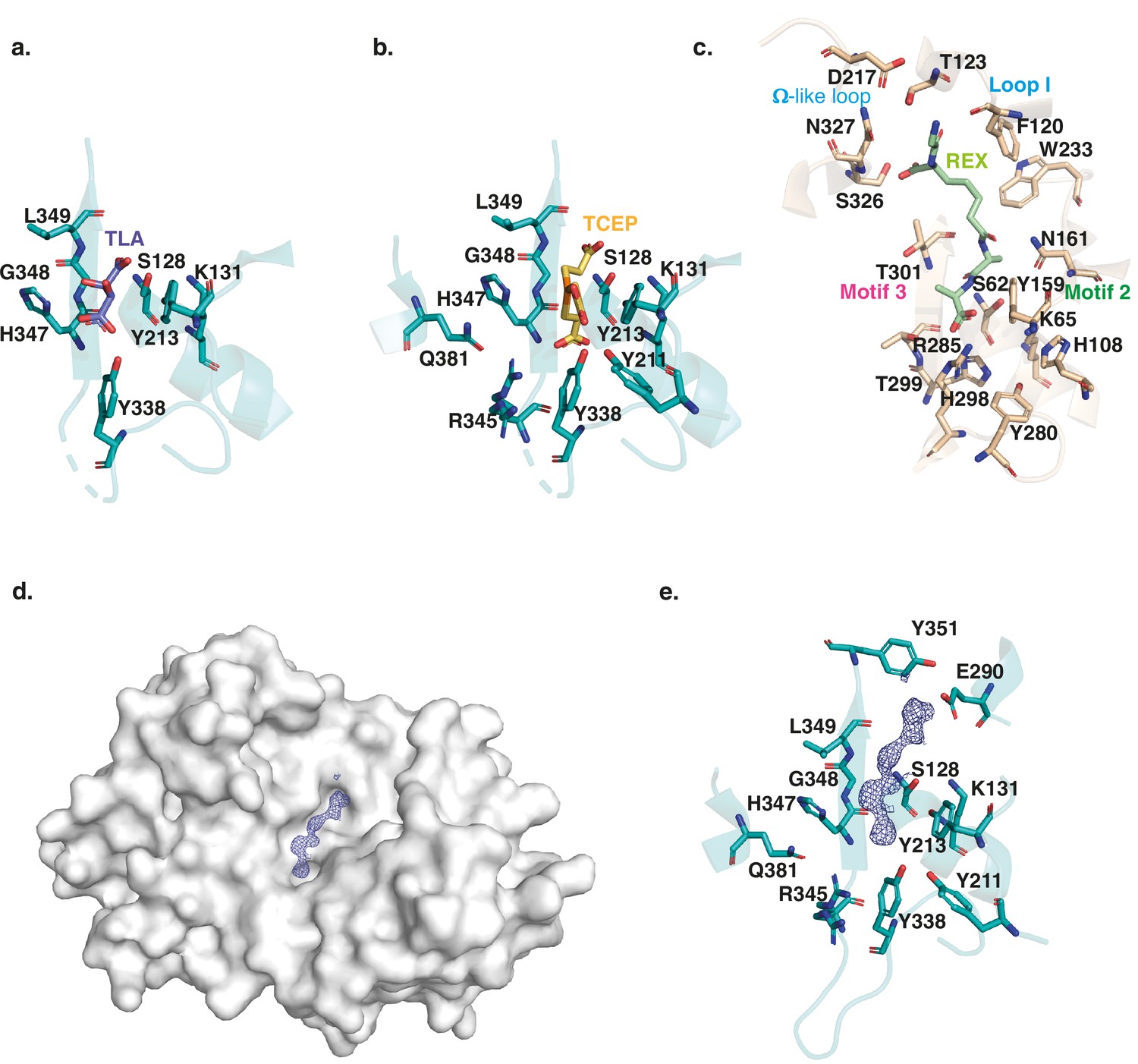
The substrate binding cleft of L. plantarum DltE.
Close-up on the ligand binding site of DltEextra crystallized in complex with a tartare (TLA) (a), or TCEP (Tris(2-carboxyethyl)phosphine hydrochloride) (b), molecule. The residues involved in the interactions are shown as sticks. The catalytic Ser128, with the hydroxyl group of its chain side is positioned at only 2.7 Å away from a carboxylic group of the ligand is itself hydrogen bonded to the catalytic Lys131 of the motif 1 and to Tyr213, the only conserved residue of the motif 2. The three residues of the β3-strand (347H-G-L349) including Gly348 from motif 3 delineate one side of the active site. Tyr338 establishes a strong, almost covalent, interaction with ligand carboxyl group that is also bound to Ser128. Three additional interactions specific to DltE are observed in the TCEP-bond structure and involved Tyr211, Arg345, and Gln381. (c) Close-up on the ligand binding site of DDCCP in complex with fragment of the cell wall precursor (REX – glycyl-L-alpha-amino-epsilon-pimelyl-D-Ala-D-Ala) (PDB ID 1IKG). The residues involved in the interactions are shown as sticks. The catalytic Ser62 and Lys65 from motif 1 and Tyr159 from motif 2 are located in the vicinity of the last D-Ala moiety in positions similar to those observed in DltE. The rest of the substrate binding site differs significantly. The Arg285 conserved in DDCP and responsible for the carboxypeptidase activity, recognizes the terminal carboxylate of the substrate. The interactions involving motif 2 and 3 stabilize the penultimate D-Ala residue. The hydrophobic subsite composed of Trp233 and Phe120 recognizes the aliphatic portion of the peptide and is also notably absent in DltE. Finally, H-bond interactions are established with the N-terminus of the peptide substrate at the level of the loop I and Ω-like-loop. (d) Surface representation of DltE structure obtained from crystal soaked with LTA molecules. A long electron density compatible with a 2-mer polyglycerol phosphate was observed lying in the catalytic cleft. (e) Close-up on the interaction network around the backbone of the ligand modeled in the catalytic cleft. The interaction network that stabilizes the first half of the ligand is the same as the one described with the TCEP molecules. The second half of the electron density extends further on the catalytic cleft of DltE in the vicinity of Glu290 (in helix α9) and Tyr351 (between β3 and β4) that are not conserved in canonical DDCP.
-
Figure 2—source data 1
DltE 3D structure in complex with tartare (TLA) or TCEP (Tris(2-carboxyethyl)phosphine hydrochloride).
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig2-data1-v2.xlsx
DltE is not active on the PG stem depsipeptide
Besides the apo form structure, we obtained two other DltE structures in complex with tartrate or TCEP (Tris(2-carboxyethyl)phosphine hydrochloride), two compounds arising from the crystallization conditions (Supplementary file 1 and Figure 2a and b). Intriguingly, these two molecules are anchored by an interaction network that involves eight different residues, among which are the conserved active site nucleophile Ser128 and the base Tyr213 (Ser 62 and Tyr 159 in DDCP; Figure 2a, b and e). Importantly, the catalytic Ser128 with the hydroxyl group of its side chain positioned at only 2.7 Å away from a carboxylic group of TCEP and tartrate is ideally placed as it would activate a substrate for the enzyme acylation event as described for DDCP (McDonough et al., 2002). This suggests that the tartrate and TCEP molecule may mimic the nature and position of the natural substrate of DltE in its active site and that the acylation/deacylation step of the catalytic reaction is probably preserved in DltE (Figure 2b and c). In addition, tartrate or TCEP established a series of interactions with the structural elements specific to DltE (see above) and not found in DDCP (Figure 2e). Notably, a Tyr residue (Tyr338 in DltE) interacting with TCEP or tartrate replaces the conserved arginine (Arg285 in DDCP) in carboxypeptidases. This arginine is key for the recognition of the terminal carboxylate of the peptide substrate and the carboxypeptidase activity (McDonough et al., 2002; Figure 2b, d and e). Furthermore, the interactions involving motif 2 and 3 that stabilizes the substrate in DDCP and in particular the penultimate D-Ala residue of the peptide substrate are also largely altered (Figure 2b, d and e). In DltE, the hydrophobic contact with TCEP or tartrate is only maintained with the conserved Tyr213 and Gly348 of motif 2 and motif 3, respectively. Instead, residues specific to DltE (Tyr338, His347, Arg345, and Leu349) stabilize the TCEP or tartrate molecules (Figure 2a and b). Taken collectively, our structural data suggest that DltE would not be able to recognize the D-Ala-D-Ala terminal end of a stem peptide as DDCP does and that DltE would not be a DD-carboxypeptidase.
In L. plantarum PG precursors, a D-Lac residue rather than the more common D-Ala is present in position 5 of the peptide stem with an ester bond between D-Ala4 and D-Lac5 (Ferain et al., 1996). Tartrate and TCEP molecules having chemical groups that are also found in D-Lac (Figure 3—figure supplement 1), we hypothesized that the DltE structural features described above could reflect its ability to act as a carboxylesterase trimming the terminal D-Lac in mature PG. However, two enzymes, namely, DacA1 and DacA2, homologous to the D-Ala-D-Ala-carboxypeptidase DacA (PBP3) of Streptococcus pneumoniae (Morlot et al., 2005) could be potentially responsible for the D-Lac hydrolysis. Thus, we generated an LpNC8 mutant deficient for dacA1 and dacA2 and analyzed the PG composition. As shown in Figure 3—figure supplement 2a, we detected the presence of disaccharide-depsipentapeptide (GlcNAc-MurNAc-L-Ala-D-Gln-mDAP-D-Ala-D-Lac). As D-Lac residues are not detected in WT strain (Bernard et al., 2011), this indicates that DacA1 and/or DacA2 could behave as carboxylesterases removing the terminal D-Lac. To confirm this, we purified the disaccharide-depsipentapeptide from the ∆dacA1∆dacA2 strain and measured the ability of the purified recombinant catalytic domains of DacA1 and DltE to catalyze D-Lac hydrolysis. We observed that DltE was not able to cleave the ester bond between D-Ala4 and D-Lac5 from the disaccharide-depsipentapeptide, whereas purified DacA1 efficiently did (Figure 3). These results indicate that DltE is not a carboxylesterase active on PG stem depsipentapeptide.
Figure 3 with 6 supplements see all
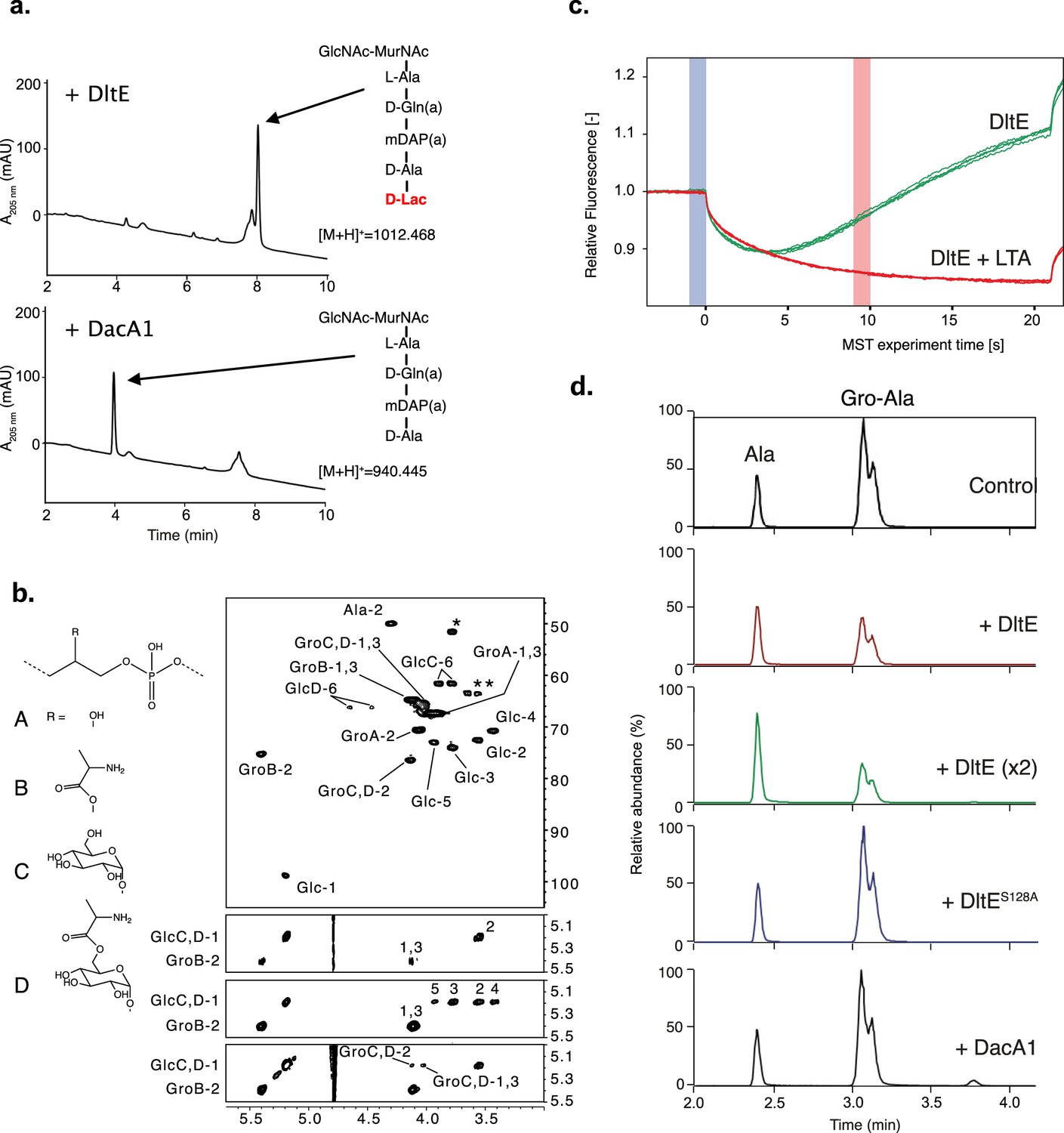
DltE is not active on peptidoglycan (PG) stem peptide but has D-Ala esterase activity on lipoteichoic acid (LTA).
(a) Test of carboxylesterase activity of DltE on disaccharide-depsipentapeptide substrate. Purified enzymes (DltEextra or DacA1) were incubated with purified muropeptide, and the mixture was analyzed by ultra-high-pressure liquid chromatography (UHPLC). DacA1 taken as a control is able to release terminal D-Lac, generating disaccharide-tetrapeptide. Muropeptides were identified by mass spectrometry. (b) Multidimensional NMR analysis of LTA isolated from WT L. plantarum established the presence of four major repeating units made of phospho-glycerol (Gro) differently substituted at C-2 position by -OH group (A), Ala- residue (B), αGlc residues (C), or Ala-6-αGlc group (D). Individual spin systems of Glc residues C and D, Gro associated to A-D and Ala associated to GroB and GlcD were established from 1H-13C HSQC (top), 1H-1H COSY (second from top), and 1H-1H TOCSY (third from top) spectra in agreement with literature (Sánchez Carballo et al., 2010). 1H and 13C chemical shifts are reported in Supplementary file 2. Ala residue was typified according to the 1H/13C chemical shifts of C1 at δ -/172.5, C2 at δ 4.29/49.9, and C3 at δ 1.63/16.36, out of which only Ala-2 is visible on the presented region of 1H-13C HSQC spectrum and Ala-1 was identified on 1H-13C HMBC spectrum (not shown). Substitution of Gro by Ala in C-2 position is established owing to the very unshielded GroB-2 signal at δ 5.40/75.3. Substitution of GroC,D in C-2 position was established owing to the deshielding of GroC,D-2 13C at δ 76.4 compared to unsubstituted GroA-2 13C at δ 70.6 and the 1H-1H NOESY cross signal between GlcC,D-1 and GroC,D-2 (bottom spectrum). Finally, substitution of GlcD in C-6 position is observable on the 1H-13C HSQC spectrum by the strong deshielding of GlcD-6 at δ 4.45–4.63/66.2 compared to unsubstituted GlcC-6 at δ 3.77–3.88/61.556. Such LTAs have never been identified in L. plantarum, but very similar repetition units were previously characterized in LTA and wall teichoic acid (WTA) isolated from Lactobacillus brevis (Sánchez Carballo et al., 2010). However, in contrast to L. brevis in which WTA and LTA showed very similar Gro-based sequences, WTA and LTA from L. plantarum showed very different structures with repetition units based either on Rbo-phosphate for the former and on Gro-phosphate for the latter. (c) Microscale thermophoresis (MST) traces for DltEextra without and with purified LTA from LpNC8 added at a concentration of 500 µM. Relative fluorescence change reveals binding of LTA to DltEextra (Fhot: fluorescence at the region defined as hot 10 s after IR laser heating, Fcold: fluorescence at the region defined as cold at 0 s). (d) Test of D-Ala esterase activity of DltE on LTA from LpNC8. Purified LTA (100 µg) was incubated with purified enzymes: DltEextra (100 µg or 200 µg [×2]), DltEextraS128A (100 µg) or DacA1 (100 µg). Control corresponds to LTA incubated in the same conditions without enzyme. After LTA depolymerization by HF treatment, hydrolysis products were analyzed by LC-MS/MS. Ala and Gro-Ala were identified by their m/z values (90.05 and 164.09, respectively) and MS-MS spectra (Figure 3—figure supplement 5). Gro-Ala was detected as a double peak, corresponding possibly to the migration of D-Ala from C2 to C1 of Gro (Morath et al., 2001). A chromatogram of the combined extracted ion chromatograms of each target ion species was generated for each condition. Similar results were obtained in two independent experiments.
-
Figure 3—source data 1
Microscale thermophoresis (MST) data for the binding of DltEextra with purified lipoteichoic acid (LTA) from LpNC8.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig3-data1-v2.xlsx
-
Figure 3—source data 2
DltE has D-Ala esterase activity on lipoteichoic acid (LTA).
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig3-data2-v2.xlsx
DltE is a D-Ala esterase acting on D-alanylated-LTA
Since DltE is not active on PG and because dltE gene is part of the dlt operon in LpNC8, we hypothesized that DltE could have a D-Ala esterase activity on D-alanylated teichoic acids. To assess whether DltE could act on WTA and/or LTA, we first characterized their structure by NMR in LpNC8 and determined their alanylation levels. Multidimensional NMR spectroscopy analysis revealed that WTA from LpNC8 contained two main repetition units composed of ribitol (Rbo)-phosphate chains substituted by two α-Glc residues either in C-2 and C-4 positions (major unit) or in C-3 and C-4 positions (minor unit) (Figure 3—figure supplement 2b). Remarkably, no Ala substituents were detected. WTAs purified from ΔdltXABCD were structurally identical from those purified from LpNC8, which supports that the DltXABCD complex is not involved in the D-alanylation of WTA. In contrast to WTA, NMR analysis established that LTA from LpNC8 is constituted by repetitive units of glycerol (Gro)-phosphate chains that were either unsubstituted (Unit A) or substituted with either Ala (Unit B), α-Glc (Unit C) or Ala-6-αGlc (Unit D) groups at C-2 position of Gro residues (Figure 3b). Relative quantification of NMR signals associated with individual units showed that Unit A was the major form with A/B/C/D ratio of 62:19:15:4. NMR analysis of LTA extracted from ΔdltXABCD mutant confirmed the absence of Ala-substituted LTA, as demonstrated by the disappearance of GroB-2 and GroB1,3 signals, GlcD-6 signals and Ala-2 signals on the 1H-13C HSQC spectrum and of GroB1 to 3 signals on the 1H-1H COSY spectrum (Figure 3—figure supplement 3). Relative quantification of signals demonstrated that LTA from ΔdltXABCD mutant consisted of a simple mixture of Unit A and C in a 70:30 ratio. Taken together, these results establish that LpNC8 cell envelope carries LTAs with a distinctive pattern of structure and substitution and that only LTAs are D-alanylated. In addition, these observations confirm that the Dlt machinery is necessary to the D-alanylation of Lp LTAs.
Given that only LTAs are alanylated in LpNC8, we started testing DltE ability to bind LTA through microscale thermophoresis (MST) experiments. As shown in Figure 3c, DltEextra was able to efficiently bind LTA. Then, we examined the activity of DltE on D-Ala-LTA purified from LpNC8. To achieve this, we analyzed the relative amounts of Ala esterified to Gro (Gro-Ala) and free Ala released from LTA upon incubation with or without DltEextra. Both compounds were detected in the control sample as a result of spontaneous release of D-Ala from LTA when the test was performed without DltE (Figure 3d and Figure 3—figure supplement 5). Upon incubation of LTA with DltE, we observed a clear decrease (56% decrease) of Gro-Ala and a concomitant increase of free D-Ala. This effect was enhanced (64% decrease) with a double amount of DltEextra in the test. In contrast, no difference with the control sample was observed for LTA incubated with DltES128A catalytic mutant protein or with DacA1 (Figure 3d and Figure 3—figure supplement 4). These results establish that DltE possesses a D-Ala esterase activity that cleaves the ester bond between the substituting D-Ala and Gro in LTA chains.
To gain structural insights into this activity, we soaked DltE crystals with LTA extracted from the ΔdltXABCD mutant and determined the 3D structure of such enzyme-substrate complexes (Supplementary file 1). We observe that the cleft harbors an additional, elongated density compatible with the presence of a 2-mer unit of the LTA molecule (Figure 2c). Half of the electron density is located at the same place as the tartrate and TCEP ligands (Figure 2) and would correspond to one unit of Gro-phosphate. It is stabilized by the same interaction network described above for tartrate and TCEP that share similar chemical groups, that is, hydroxyl, carboxyl and phosphate with LTA (Figure 2d, Figure 3—figure supplement 1). The second half of the electron density extends further on the catalytic cleft of DltE in the vicinity of Glu290 (in helix α9) and Tyr351 (between β3 and β4), forming thus a negatively charged subsite. The latter is not conserved in DDCP and replaced by a hydrophobic subsite that was proposed to recognize the aliphatic portion of the peptide (Figure 3—figure supplement 6a and b). These structural features are in agreement with the ability of DltE to bind and hydrolyse D-Ala-LTA, and, together with the biochemical data, they confirm that DltE is a D-Ala esterase acting on D-Ala-LTA.
The D-Ala esterase activity of DltE contributes to D-alanylation of the cell envelope and is required to sustain Drosophila juvenile growth
We previously reported that the machinery encoded by the dlt operon (including dltE) is necessary to support D-Ala esterification in Lp cell envelope and sustain Drosophila juvenile growth (Matos et al., 2017). As previously observed upon mild alkaline hydrolysis, D-Ala was released in appreciable amounts from LpNC8 (WT) cells14 (Figure 4a), whereas no D-Ala was released from an isogenic mutant deleted for the entire dlt operon (Matos et al., 2017). However, we observed a significant reduction (around 70%) of D-Ala esterified to LTAs in ΔdltE mutant cells and an even higher reduction (around 93%) for DltES128A catalytic mutant cells. These results indicate that the esterase activity of DltE, in addition to the activities of DltX, A, B, C, and D, contributes to the D-alanylation machinery of LTAs in bacterial cells.
Figure 4 with 1 supplement see all
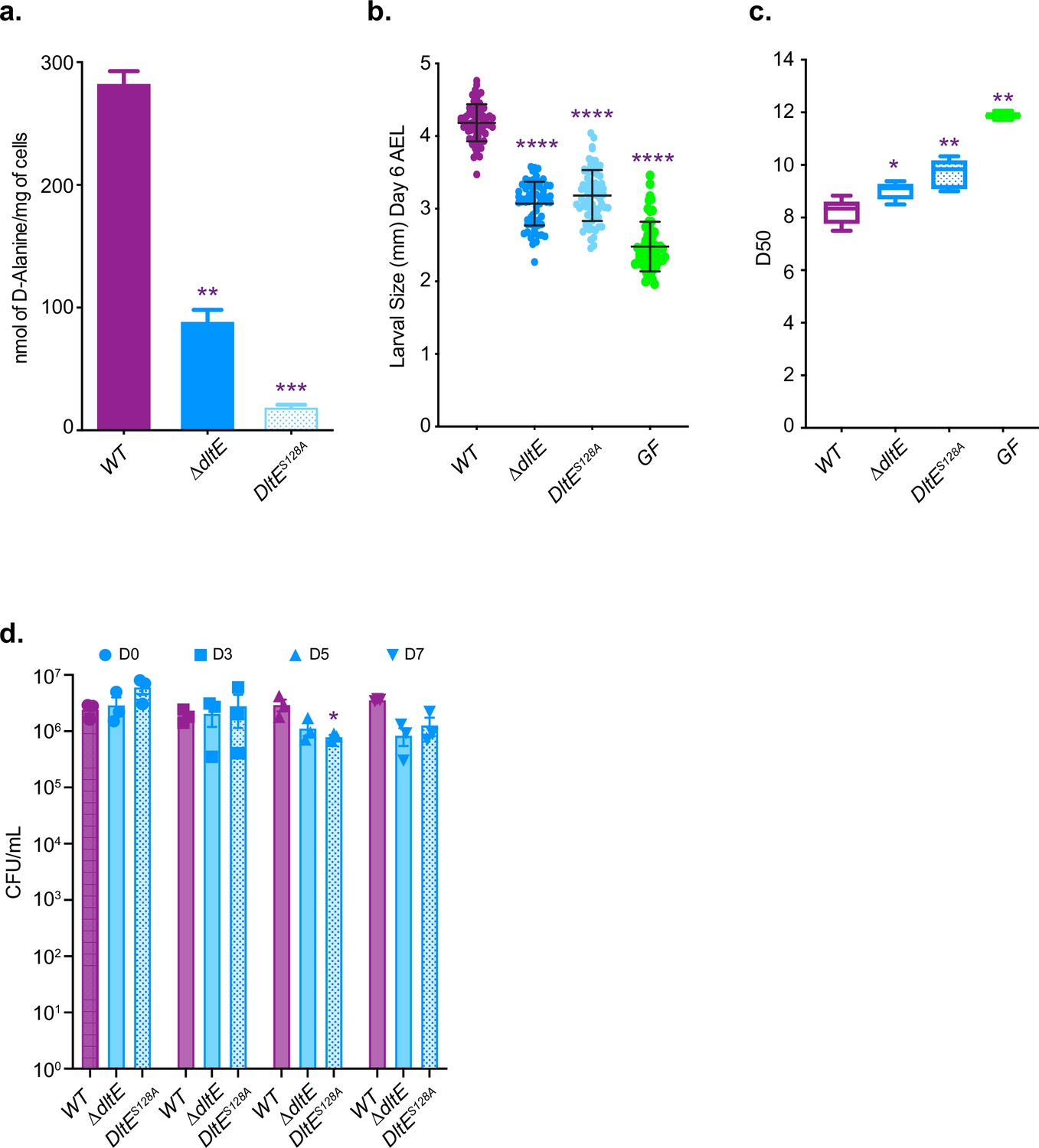
The D-Ala esterase activity of DltE contributes to D-alanylation of the cell envelope and is required to sustain Drosophila juvenile growth.
(a) Amount of D-Ala released from whole cells of WT, ∆dltE, and DltES128A by alkaline hydrolysis and quantified by HPLC. Mean values were obtained from three independent cultures with two injections for each. Purple asterisks illustrate statistically significant difference with D-Ala release from WT. ***0.0001<p<0.001; **0.001<p<0.01. (b) Larval longitudinal length after inoculation with WT, ∆dltE, and DltES128A strains and PBS (for the germ-free [GF] condition). Larvae were collected 6 d after association and measured as described in the ‘Materials and methods’ section. Purple asterisks illustrate statistically significant difference with larval size of WT; ****p<0.0001. Center values in the graph represent means and error bars represent SD. Representative graph from one out of three independent experiments. (c) Time to pupation: day when 50% of pupae emerge during a developmental experiment (D50) for GF eggs associated with strains WT, ∆dltE, and DltES128A or PBS (for the GF condition). Center values in the graph represent means. Purple asterisks illustrate statistically significant difference with WT larval size; **0.001<p<0.01; *p<0.05. (d) Abundance of colony-forming units (CFUs) on fly food and larvae at days 3, 5, and 7 after inoculation with WT, ∆dltE, and DltES128A. Purple asterisks illustrate statistically significant difference with WT within each day. *p<0.05.
-
Figure 4—source data 1
The D-Ala esterase activity of DltE contributes to D-alanylation of the cell envelope and is required to sustain Drosophila juvenile growth.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig4-data1-v2.xlsx
-
Figure 4—source data 2
The D-Ala esterase activity of DltE is required to sustain Drosophila juvenile growth – replicates.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig4-data2-v2.xlsx
We next wondered whether an active DltE protein is necessary for L. plantarum support to Drosophila juvenile growth. To this end, we compared juvenile growth (larval size at day 6 after egg laying) and developmental timing (i.e. day of 50% population entry to metamorphosis) of germ-free animals and ex-germ-free animals associated at the end of embryogenesis with either WT or ΔdltE and dltES128A mutants. Both mutations did not affect the ability of Lp to thrive in the fly niche but significantly altered the ability of Lp to support larval growth and developmental timing (Figure 4b–d and Figure 4—figure supplement 1). These results demonstrate that DltE activity is required to sustain Drosophila juvenile growth and together with the structural and biochemical insights on DltE activity point to the importance of D-Ala-LTAs as cues supporting Drosophila growth.
D-Ala-LTAs are necessary bacterial cues supporting Drosophila intestinal response and juvenile growth
In order to evaluate the importance of D-Ala-LTA for Drosophila juvenile growth, we undertook a genetic approach by generating L. plantarum strains either deprived of LTA through the deletion of ltaS encoding for the LTA synthetase (ΔltaS) (Gründling and Schneewind, 2007) or WTA through the deletion of tagO encoding for the enzyme catalyzing the first step of WTA biosynthesis (ΔtagO) (Andre et al., 2011). A similar amount of D-Ala esterified to the cell envelope was measured in ΔtagO mutant cells compared to WT cells, whereas a strong reduction of D-Ala (around 72%) was observed in ΔltaS mutant cells (Figure 5—figure supplement 1a). The absence of LTA chain synthesis in ΔltaS mutant was confirmed by western blot with anti-LTA monoclonal antibody (Figure 5—figure supplement 1b). These results confirm that LTA are D-alanylated but not WTA as determined above by NMR analysis of purified polymers (Figure 3). Of note, we hypothesize that the residual D-Ala released for ΔltaS cells might be attached to a glycolipid precursor of LTA as previously observed in Listeria monocytogenes (Webb et al., 2009).
We then tested the respective TA defective strains for their ability to support Drosophila juvenile growth (Figure 5a and Figure 5—figure supplement 2a). When associated with germ-free animals, WT and ΔtagO strains support optimal Drosophila juvenile growth while Δdltop or ΔltaS strains largely fail to do so despite colonizing well their host’s niche (Figure 5a, Figure 5—figure supplement 1c, and Figure 5—figure supplement 2a). These data demonstrate that LTA are important molecules mediating Lp’s support to Drosophila juvenile growth. Accordingly, presence of D-Ala esterified to LTA chains (in WT and ΔtagO strains) correlates well with strains ability to support Drosophila’s growth. Indeed, D-Ala-LTA presence is required for Lp-mediated Drosophila juvenile growth promotion phenotype (Figure 5a, Figure 5—figure supplement 1a and b, and Figure 5—figure supplement 2a). In the absence of D-Ala-LTA (in Δdltop) or LTA (in ΔltaS), Drosophila larvae are smaller (Figure 5a and Figure 5—figure supplement 2a). Taken collectively these data reinforce the notion that D-Ala-LTAs from Lp play a key role in supporting Drosophila juvenile growth.
Figure 5 with 2 supplements see all
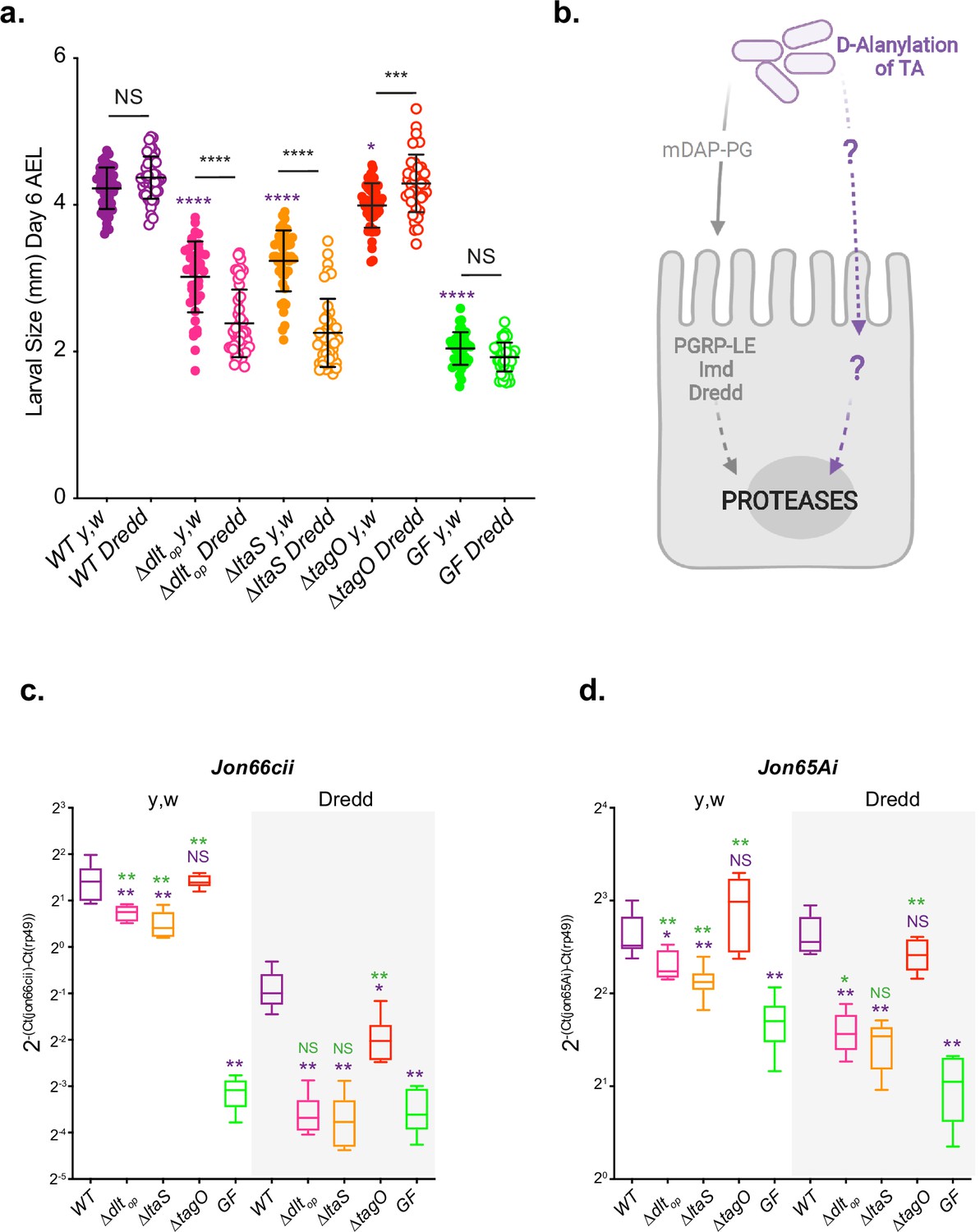
D-Ala-LTAs are necessary bacterial cues supporting Drosophila intestinal response and juvenile growth.
(a) y,w and y,w,Dredd larvae longitudinal length after inoculation with 108 colony-forming units (CFUs) of WT, ∆dltop, ∆ltaS, and PBS, for the germ-free condition. Larvae were collected 6 d after association and measured as described in the ‘Materials and methods’ section. The purple asterisks represent a statistically significant difference compared with WT larval size. The bars in the graph represent means and SD. NS represents the absence of a statistically significant difference; ****p<0.000; ***0.0001<p<0.001; *p<0.05. Representative graph from one out of three independent experiments. (b) Working model for Lp detection in Drosophila enterocytes: Drosophila sense and signal the presence of Lp cells through: (1) PGRP-LE-mediated mDAP-PG fragment recognition triggering Imd/Dredd signaling and (2) sensing of bacterial cell envelope bearing D-alanylated teichoic acids and signaling by unknown host mechanisms. Both signals were reported to be important for maximal intestinal peptidase expression. (c, d) Mean ± SD of 2-∆Ctgene/∆Ctrp49 ratios for Jon66Cii and Jon65Ai detected in dissected guts of y,w and y,w,Dredd associated with WT, ∆dltop, ∆ltaS or the GF condition from five biological replicates. Representative graphs from one out of three independent experiments are shown. The purple asterisks represent a statistically significant difference compared with LpNC8 proteases expression. The green asterisks represent a statistically significant difference compared with the GF condition. NS represents the absence of a statistically significant difference compared to the GF condition or LpNC8. **0.001<p<0.01; *p<0.05.
-
Figure 5—source data 1
D-Ala-LTAs are necessary bacterial cues supporting Drosophila intestinal response and juvenile growth.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig5-data1-v2.xlsx
-
Figure 5—source data 2
D-Ala quantification and persistence on fly niche of WT, ∆dltop, ∆ltaS, and ∆tagO strains.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig5-data2-v2.xlsx
-
Figure 5—source data 3
D-Ala-LTAs are necessary bacterial cues supporting Drosophila intestinal response and juvenile growth – replicates.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig5-data3-v2.xlsx
-
Figure 5—source data 4
Replicate western blot detection of lipoteichoic acid (LTA) in wild-type LpNC8 and mutant derivatives.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig5-data4-v2.pdf
We had previously established that upon association with Lp, enterocytes from ex-germ-free Drosophila sense and signal the presence of Lp cells by at least two independent molecular mechanisms: (1) PGRP-LE-mediated mDAP-PG fragment recognition triggering Imd/Dredd signaling (Erkosar et al., 2015) and (2) sensing of bacterial cell envelope bearing D-alanylated teichoic acids and signaling by unknown host mechanisms (Figure 5b; Matos et al., 2017). Both signals were reported to be important for maximal intestinal peptidase expression and optimal support to growth promotion by Lp (Figure 5a, c and d and Figure 5—figure supplement 2a–c; Matos et al., 2017). To probe the respective contribution of LTA and WTA to this model, we tested the ability of ΔltaS and ΔtagO strains (respectively deprived of LTA or WTA) to support Drosophila growth and intestinal peptidase expression in a Drosophila genetic background (Dredd mutants flies) (Leulier et al., 2000) that blunts the host response to mDAP-PG fragments (Leulier et al., 2003), allowing us to focus on the contribution of the D-alanylation signal to Lp-mediated growth support and intestinal peptidase induction. As previously observed upon association with the Δdltop mutant, Dredd mutant larvae associated with ΔltaS are compromised in their juvenile growth potential (Figure 5a and Figure 5—figure supplement 2a) and their ability to stimulate intestinal peptidase expression (Jon66Cii and Jon65Ai; Figure 5c and d and Figure 5—figure supplement 2b and c). However, juvenile growth and intestinal peptidase expression were not markedly affected in ΔtagO-associated Dredd larvae (Figure 5a–d and Figure 5—figure supplement 2a–c). Importantly, we observed a cumulative effect of not having D-Ala-LTA (Δdltop) or not having LTA at all (ΔltaS) and altering host mDAP-PG sensing (compare WT -yw- and Dredd conditions, Figure 5a–d and Figure 5—figure supplement 2a–c). These results therefore support the notion that both signals are required for optimal support to larval growth by Lp and that in addition to mDAP-PG fragments, D-Ala-LTAs are likely an additional cue sensed by Drosophila enterocytes.
D-Ala-LTAs are direct bacterial cues supporting Drosophila intestinal response and juvenile growth
Next, we tested the hypothesis that D-Ala-LTAs act as direct signals sensed by Drosophila enterocytes. To this end, we purified the major components from the cell envelope of WT and ΔdltXABCD strains: WTA, LTA, and PG, and tested their ability to rescue Δdltop and ΔltaS-mediated larval phenotypes (Figure 6a–e and Figure 6—figure supplement 1a–d). First, germ-free, Δdltop or WT-associated animals were treated daily with the purified cell envelope components (WTA, PG, and LTA) for 5 d. On day 6, larvae were harvested and their size measured (Figure 6b–e and Figure 6—figure supplement 1a–d). Daily supplementation with purified WTA or PG either from WT or ΔdltXABCD strains did not impact the growth of GF or Δdltop-associated larvae (Figure 6b and c and Figure 6—figure supplement 1a and b). In contrast, the daily supplementation of purified LTA isolated from WT (D-Ala-LTAs) to larvae associated with the Δdltop strain shows a growth-promoting effect that leads to increased larval size in this condition compared to the non-supplemented control (Figure 6d and e). This growth-promoting effect was not observed in GF and WT-associated animals nor when Δdltop associated larvae were supplemented with non-alanylated-LTAs isolated from ΔdltXABCD strain (Figure 6d and e and Figure 6—figure supplement 1c and d). We repeated the similar experiment with ΔltaS-associated larvae and again observed an improved larval growth upon supplementation with LTA purified from WT (D-Ala-LTAs) but not from LTAs purified from ΔdltXABCD strain (i.e. non-alanylated-LTAs) (Figure 6e and Figure 6—figure supplement 1d). These results therefore establish that D-Ala-LTAs are necessary and sufficient to restore Drosophila juvenile growth.
Figure 6 with 2 supplements see all
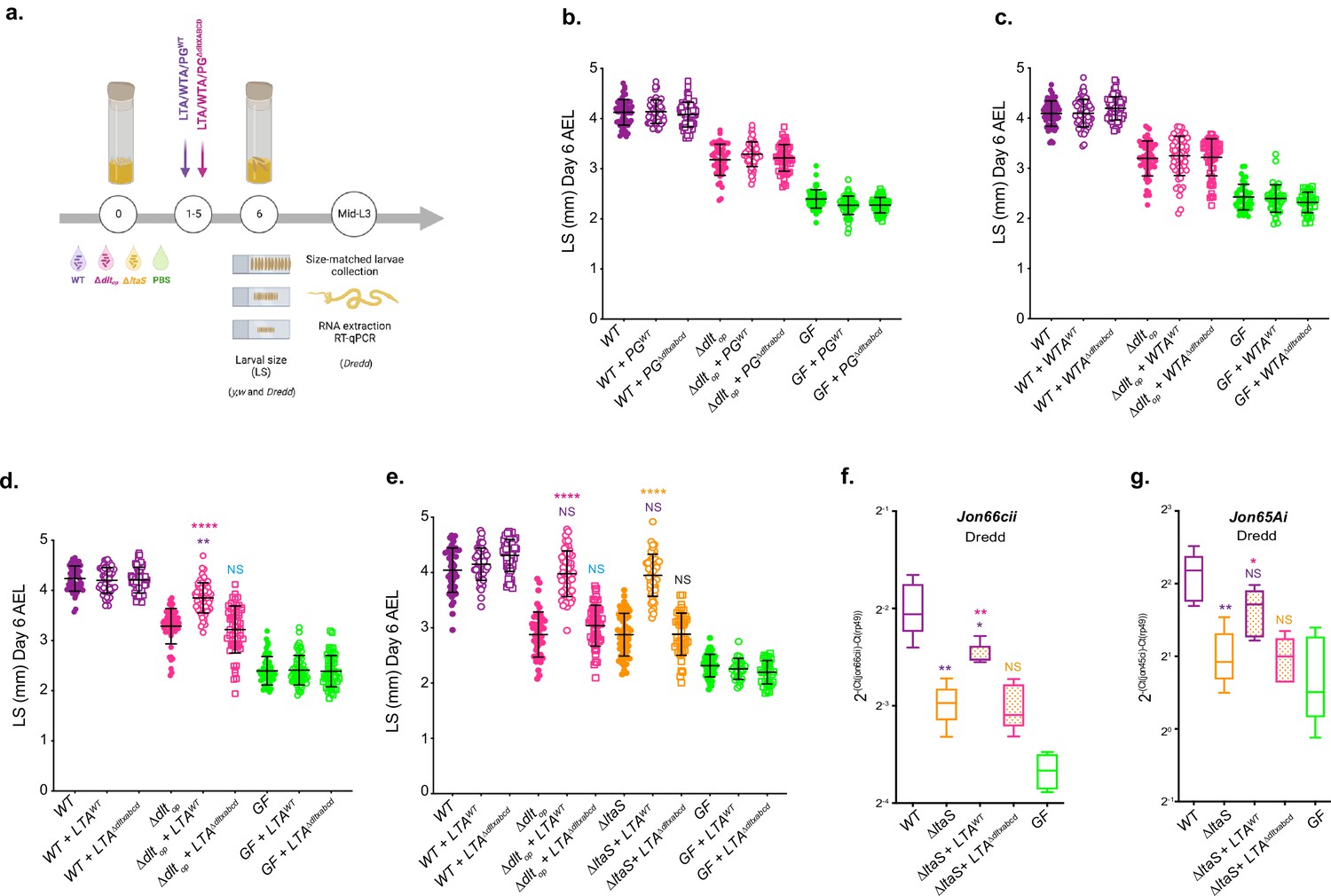
D-Ala lipoteichoic acids (LTAs) are bacterial cues supporting Drosophila intestinal response and juvenile growth.
(a) Experimental set-up to test the impact of cell envelope components on Drosophila growth and proteases expression: germ-free (GF) eggs were inoculated with WT, ∆dltop, ∆ltaS, or PBS for the GF condition and supplemented daily (for 5 d) with LTA, wall teichoic acid (WTA), and peptidoglycan (PG) extracted from WT or ∆dltXABCD strains. 1 µg of each component purified from WT was used. For comparison with components extracted ΔdltXABCD strain, the final PG, WTA, and LTA suspensions were adjusted to the same amount of Gro, Rbo, and Mur, respectively. At day 6 after inoculation, larvae were harvested and measured (see the ‘Materials and methods’ section for details). Mid-L3 sized-matched larvae were collected for each condition. Their guts were dissected followed by RNA extraction and RT-qPCR targeting proteases expression. (b–d) Larval longitudinal length after inoculation with strains WT, ∆dltop, PBS, and purified PG (PG) (b) WTA (c) or LTA (d) from WT or ∆dltXABCD. Larvae were collected 6 d after the first association and measured as described in the ‘Materials and methods’ section. The pink asterisks represent a statistically significant difference compared with ∆dltop larval size; purple asterisks represent a statistically significant difference compared with WT larval size. NS represents the absence of a statistically significant difference compared to ∆dltop. ****p<0.0001; **0.001<p<0.01. The bars in the graph represent means and SD. A representative graph from one out of three independent experiments is shown. (e) Larval longitudinal length after inoculation with strains WT, ∆dltop, ∆ltaS, or PBS and purified LTA from WT or ∆dltXABCD strains. Larvae were collected 6 d after the first association and measured as described in the ‘Materials and methods’ section. The pink asterisks represent a statistically significant difference compared with ∆dltop larval size; purple asterisks represent a statistically significant difference compared with WT larval size ****p<0.0001; **0.001<p<0.01. The bars in the graph represent means and SD. A representative graph from one out of three independent experiments is shown. (f, g), Mean ± SD of 2-∆Ctgene/∆Ctrp49 ratios for Jon66Cii and Jon65Ai detected in dissected guts of y,w,Dredd size-matched larvae, associated with LpNC8, ∆dltop, ∆ltaS or the GF and LTA from WT or ∆dltXABCD strains, from five biological replicates. Representative graphs from one out of three independent experiments are shown. The purple asterisks represent a statistically significant difference compared with LpNC8 protease expression. The pink asterisks represent a statistically significant difference compared with ∆ltaS supplemented with LTA from ∆dltXABCD strain. NS represents the absence of a statistically significant difference compared to ∆ltaS condition. **0.001<p<0.01; *p<0.05.
-
Figure 6—source data 1
D-Ala lipoteichoic acids (LTAs) are bacterial cues supporting Drosophila intestinal response and juvenile growth.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig6-data1-v2.xlsx
-
Figure 6—source data 2
Experimental replicates from Figure 6b–d.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig6-data2-v2.xlsx
-
Figure 6—source data 3
Experimental replicates from Figure 6f and g.
- https://cdn.elifesciences.org/articles/84669/elife-84669-fig6-data3-v2.xlsx
Finally, we repeated purified LTA supplementations on Dredd larvae associated with the ΔltaS strain for 5 d and dissected the midguts of size-matched larvae to analyze intestinal peptidase (Jon66cii and Jon65Ai) expression by RT-qPCR independently of any mDAP-PG sensing/Imd signaling input (Figure 6a, f and g and Figure 6—figure supplement 2). We observed that supplementation with D-Ala-LTAs was sufficient to recapitulate the effect of WT strain on intestinal peptidase expression even in the absence of any mDAP-PG signal (Figure 5b, Figure 6f and g, and Figure 6—figure supplement 2). Taken collectively, our results demonstrate that in addition to and independently of mDAP-PG signaling, D-Ala-LTAs are important direct bacterial cues supporting intestinal peptidase expression and Drosophila juvenile growth.
Discussion
The results obtained in this study pinpoint the bacterial machinery and molecular determinant of the bacterial cell envelope involved in the beneficial relationship between Drosophila and selected strains of its commensal partner Lp, which results in optimized host growth. Previously, we identified through forward genetic screening the pbpX2-dltXABCD operon encoding a bacterial machinery essential to LpNC8-mediated support to Drosophila larval growth. Here, using complementary structural, biochemical, and genetic approaches, we reveal that the protein encoded by the first gene of this operon (now renamed DltE) is not a bona fide PBP as it does not bind penicillin nor modifies Lp PG. Instead, we establish that DltE is a D-alanine esterase acting upon LTA. While studying its physiological function, we discovered that DltE contributes together with the other proteins encoded by the dlt operon (the DltX-A-B-C-D machinery) to the alanylation of LTA on its glycerol residues. The study of DltE’s substrates gave insights on the structure and composition of LpNC8’s TAs, which revealed that only LTA and not WTA are alanylated in this strain. Finally, prompted by this strain-specific feature and the importance of the dltEXABCD operon for LpNC8-mediated support to Drosophila growth, we identified that LTAs alanylated by the DltE/X/A/B/C/D complex (but not WTA, which are not alanylated, nor non-alanylated LTA) act as direct bacterial clues supporting Drosophila intestinal function and juvenile growth.
DltE and the Dlt machinery
Regardless of the initial DltE in silico annotation as a PBP (PbpX2), our structural data revealed that the global architecture of its active site cavity is reshaped and different from that of canonical PBP enzymes suggesting an alternative substrate recognition mode and/or a different substrate specificity. Hence, we show that D-Ala LTA and not PG is the substrate of DltE. Given its atomic structure and D-Ala esterase activity, DltE may rather relate to Staphylococcus aureus FmtA, a D-amino esterase acting on teichoic acids (Rahman et al., 2016). FmtA is, however, different from DltE given that it shows residual PBP activity, is able to interact with ß-lactams, and is encoded apart from the dltABCD operon in S. aureus genome. So far, the hypothesis advanced for FmtA biological function concerns the removal of D-Ala from LTAs to make it available to transfer onto WTAs or removal of D-Ala from WTA to reset cell surface charge under particular conditions (Rahman et al., 2016). Considering that WTA are not alanylated in LpNC8, these observations suggest that DltE possesses a distinct function than FmtA. We notice an apparent discrepancy between DltE enzymatic activity and the D-alanine content of the dltE mutants (ΔdltE and dltES128A). We posit that DltE may be involved in the control of D-Ala distribution on the LTA chains and DltE inactivation would result in impaired functioning of the whole D-alanylation machinery. Our work paves the way to the characterization of the complete mechanistical framework in which the interplay between the Dlt machinery and DltE are required for the D-alanylation of LTAs.
Strain specificity in L. plantarum teichoic acids composition and modifications
We analyzed LpNC8 TAs composition through multidimensional NMR spectroscopy and determined their alanylation levels. LpNC8 LTAs are, as expected from this strain’s genomic information and previous Gram-positive bacteria LTA characterization (Percy and Gründling, 2014; Sutcliffe, 2011), constituted by poly-(Gro-phosphate) chains substituted, among others, with Ala. In contrast, but in agreement with the genetic information from LpNC8 genome, this strain produces poly(Rbo-phosphate) WTAs given the lack of the tag genes encoding the machinery responsible for the synthesis of poly-(Gro-phosphate) chains in its genome (Bron et al., 2012). The ability to produce poly-(Gro-phosphate) WTA chains is a strain-specific feature in Lp as the tag gene cluster is only found in 22 out of 54 strains recently studied at the genomic level (Martino et al., 2016). However, in the case of the LpWCFS1 strain, despite its genome carrying the genetic information to synthetize both types of WTA (poly-(Gro-phosphate) and poly-(Rbo-phosphate)), only poly(Gro-phosphate) WTAs were detected at steady state (Bron et al., 2012). Surprisingly, no Ala substituents were detected in LpNC8 WTAs. This is an original feature as in other L. plantarum strains such as LpWCFS1, WTA appears to be D-alanylated (Bron et al., 2012). Yet, these WTAs are composed of poly(Gro-phosphate) chains so it would be interesting to investigate the levels of D-alanylation in other Lp strains harboring only poly(Rbo-phosphate) WTAs. Taken together, these observations call for future systematic analysis of TA composition and modification among L. plantarum strains and question the potential correlation between WTA composition and/or TA D-alanylation patterns and the ability of the strain to benefit Drosophila growth.
Drosophila response to commensals’ cell envelope determinants
We have previously established that PG fragments (Erkosar et al., 2015) and D-Ala esterification to the cell envelope (Matos et al., 2017) trigger intestinal peptidase expression and support Drosophila juvenile growth. Here, we report that in addition to PG fragments, D-Ala-LTA from LpNC8 cell envelop represent a bacterial cue necessary and sufficient for the ability of LpNC8 to promote Drosophila growth and upregulation of intestinal digestive peptidases in enterocytes. Given that L. plantarum is a gut luminal microbe and that the Drosophila midgut is lined with both thick chitinous matrix and thin mucus layer, yet permeable to large macromolecules and small nutrient particles (Lemaitre and Miguel-Aliaga, 2013), how exactly these macromolecules associated to the bacterial cell membrane reach the enterocytes remains elusive. Similarly to PG fragments, which are shaded in the extracellular media during bacterial cell division, LTA chains may also be shed after cleavage of the lipid anchor (Neuhaus and Baddiley, 2003). In addition, our recent findings report LpNC8’s ability to release microvesicles of multi-nanometers ranges (Grenier et al., 2021). This observation is coupled to previous reports showing that other Lactobacillus strains form microvesicles containing LTAs that are endocytosed by mammalian enterocytes (Champagne-Jorgensen et al., 2021), it is tempting to speculate that LpNC8 D-Ala-LTAs are released via bacterial microvesicles cargo. The Drosophila host would capitalize on these features by endocytosing cleaved LTA or such cargo and sense these bacterial cues through pattern recognition receptors expressed in enterocytes.
PG fragments are sensed intracellularly in the Drosophila midgut by PGRP-LE, which signals the Imd pathway to stimulate intestinal peptidase expression. How D-Ala-LTAs are sensed and signal in enterocytes remain elusive, but our results support the notion that another signaling cascade beyond the one engaged by PGRP-LE is leading to D-Ala-LTA-mediated intestinal peptidase induction. Taken together, it seems that Drosophila ensures an optimal intestinal response to bacterial cues by using additive rather than interdependent signals converging locally to intestinal peptidase expression and macroscopically to enhanced juvenile growth.
Given the strong analogies of the impact of L. plantarum strains on Drosophila and mouse growth (Schwarzer et al., 2016) and previous work focusing on LTA in host-Gram-positive bacteria interactions (Champagne-Jorgensen et al., 2021; Hara et al., 2018), an exciting perspective will be to test the contribution of D-Ala-LTA to L. plantarum-mediated host growth promotion in other symbiotic models including mammals.
Materials and methods
Drosophila diets, stocks and breeding
Request a detailed protocolDrosophila stocks were cultured as described in Erkosar et al., 2015. Briefly, flies were kept at 25°C with 12/12 hr dark/light cycles on a yeast/cornmeal medium containing 50 g/L of inactivated yeast. The low-yeast diet was obtained by reducing the amount of inactivated yeast to 7 g/L. Germ-free stocks were established as described in Erkosar et al., 2015. Axenicity was routinely tested by plating serial dilutions of animal lysates on nutrient agar plates. Drosophila y,w flies were used as the reference strain in this work. The following Drosophila line was also used: y,w,DreddF64 (Leulier et al., 2000).
Bacterial strains and growth conditions
Request a detailed protocolStrains used in this study are listed in Supplementary file 4. Escherichia coli strains were grown at 37°C in LB medium with agitation. L. plantarum strains were grown in static conditions in MRS media at 37°C, unless differently stated. Erythromycin antibiotic was used at 5 μg/mL for L. plantarum and 150 μg/mL for E. coli in the context of the deletion strains construction.
Construction of deletion strains in L. plantarumNC8
Request a detailed protocolIndependent markerless deletions on ltaS, tagO, dacA1, and dacA2 genes of LpNC8 genome were constructed through homology-based recombination with double-crossing over as described by Matos et al., 2017 . Briefly, the 5′- and 3′-terminal regions of each region were PCR-amplified with Q5 High-Fidelity 2X Master Mix (NEB) from LpNC8 chromosomal DNA. Primers contained overlapping regions with pG+host9 (Maguin et al., 1996) to allow for Gibson Assembly. PCR amplifications were made using the following primers: XL01/XL02 and XL03/XL04 (ltaS), XL05/XL06 and XL07/XL08 (tagO), XL09/XL10 and XL11/XL12 (dacA1), XL13/XL14, and XL15/XL16 (dacA2) listed in Supplementary file 3. The resulting plasmids obtained by Gibson Assembly (NEB) were transformed into LpNC8 electrocompetent cells and selected at the permissive temperature (28°C) on MRS plates supplemented with 5 µg/mL of erythromycin. Overnight cultures grown under the same conditions were diluted and shifted to the non-permissive temperature (41°C) in the presence of 5 µg/mL of erythromycin to select single crossover integrants. Plasmid excision by a second recombination event was promoted by growing integrants at the permissive temperature without erythromycin. Deletions were confirmed by PCR followed by sequencing. The strain deleted for dacA1 and dacA2 was obtained by the sequential deletion of dacA1 followed by dacA2.
Knock-in of DltE catalytic dead versions in L. plantarumNC8
Request a detailed protocolL. plantarumNC8 strain carrying a modified version of the dltE gene (dltES128A) was built by knocking in the modified sequence on ΔdltE strain (Matos et al., 2017). dltE-modified sequence harboring the mutation S128A was obtained by PCR with modified primers (X19). The 5′- and 3′-terminal regions of dltE region together with the entire dltE gene were PCR-amplified with Q5 High-Fidelity 2X Master Mix (NEB) from L. plantarumNC8 chromosomal DNA using primers XL17/XL18 and XL19/XL20 (Supplementary file 3). The two fragments were assembled with pG+host9 (Maguin et al., 1996). The resulting plasmid was transformed into ΔdltE electrocompetent cells and selected at the permissive temperature (28°C) on MRS plates supplemented with 5 µg/mL of erythromycin. Overnight cultures grown under the same conditions were diluted and shifted to the non-permissive temperature (41°C) in the presence of 5 µg/mL of erythromycin to select single crossover integrants. Plasmid excision by a second recombination event was promoted by growing integrants at the permissive temperature without erythromycin. dltE S128A knock-in was confirmed by PCR followed by sequencing.
Larval size measurements
Request a detailed protocolAxenic adults were put overnight in breeding cages to lay eggs on sterile low-yeast diet. Fresh axenic embryos were collected the next morning and seeded by pools of 40 in tubes containing fly food. 1 × 108 CFUs or PBS were then inoculated homogeneously on the substrate and the eggs. Fly tubes are incubated at 25°C until larvae collection. Drosophila larvae, 6 d after inoculation, were randomly collected and processed as described by Erkosar et al., 2015 . Individual larval longitudinal length was quantified using ImageJ software (Schneider et al., 2012). For the cell envelope components supplementation experiments, 1 µg of purified LTA, WTA, and PG (extracted from strains LpNC8 and ΔdltXABCD) resuspended in PBS were added, independently, to the fly food every day until day 5 (see Figure 6a). For comparison of components extracted from each of the two strains, the final LTA, WTA, and PG suspensions were adjusted to the same amount of Gro, Rbo, and Mur, respectively. At day 6, larvae were harvested and larval longitudinal length was quantified using ImageJ software (Schneider et al., 2012).
Developmental timing determination
Request a detailed protocolAxenic adults were placed in breeding cages overnight to lay eggs on sterile poor-yeast diet. Fresh axenic embryos were collected the next morning and seeded by pools of 40 in tubes containing fly food. A total of 1 × 108 CFUs of each strain or PBS was then inoculated homogeneously on the substrate and the eggs and incubated at 25°C. The emergence of pupae was scored every day until all pupae had emerged. D50 (day when 50% of the pupae emerged) was determined using D50App (Matos et al., 2017).
Bacterial loads analysis
Request a detailed protocolTo access bacterial CFU in the fly nutritional matrix, microtubes containing food and larvae were inoculated with 1 × 107 CFUs of each strain, independently. Tubes were incubated at 25°C until being processed. For bacterial load quantification, 0.75–1 mm glass microbeads and 500 μL PBS were added directly into the microtubes. Samples were homogenized with the Precellys 24 tissue homogenizer (Bertin Technologies). Lysates dilutions (in PBS) were plated on MRS agar using the Easyspiral automatic plater (Intersciences). MRS agar plates were then incubated for 24 hr at 37°C. Bacterial concentration was deduced from CFU counts on MRS agar plates using the automatic colony counter Scan1200 (Intersciences) and its counting software. For larval loads, y,w axenic eggs were inoculated with 1 × 108 CFUs of each strain and incubated at 25°C until collection. Size-matched larvae were harvested from the nutritive substrate and surface-sterilized with a 30 s bath in 70% ethanol under agitation and rinsed in sterile water. Pools of five larvae were deposited in 1.5 mL microtubes containing 0.75–1 mm glass microbeads and 500 μL of PBS.
RNA extraction and RT-qPCR analysis
Request a detailed protocolAxenic y,w and y,w,Dredd embryos were inoculated with 1 × 108 CFU of LpNC8, Δdltop, and ΔltaS strains independently or kept axenic. Larvae were size matched for the four conditions and harvested at mid-L3 larval stage. Alternatively, larvae were inoculated with each one of the strains mentioned above and supplemented daily with cell envelop purified components (LTA, WTA, PGN) from LpNC8 or Δdltop cells. RNA extraction of five replicates of six dissected guts for each condition was performed as described by Matos et al., 2017. RT-qPCR was performed using gene-specific primer sets (Supplementary file 3) as described by Matos et al., 2017. Results are presented as the value of ∆Ctgene/∆Ctrp49.
Statistical analysis
Request a detailed protocolData representation and analysis was performed using GraphPad PRISM 8 software (https://www.graphpad.com). A total of 3–5 biological replicates were used for all experiments performed in this study in order to ensure representativity and statistical significance. All samples were included in the analysis. Experiments were done without blinding. Two-sided Mann–Whitney’s test was applied to perform pairwise statistical analyses between conditions.
E. coli plasmid construction
Request a detailed protocolDNA fragments were amplified by polymerase chain reaction using L. plantarum cDNA as a template and oligonucleotides listed in Supplementary file 5. The DNA encoding the extracellular domain of wild-type DltE (DltEextra) and the catalytic mutant DltEextra-S128A were cloned into the NcoI and XhoI sites of the pET-28a(+) vector that expresses proteins fused to a C-terminal hexahistidine (His)6 tag (Supplementary file 6).
Protein production and purification
Request a detailed protocolDltEextra and DltEextra-S128A were expressed in E. coli BL21 (DE3)-RIPL cells. Cells were grown in LB media at 37°C and induced with 0.5 mM isopropyl b-D-1-thiogalactopyranoside (IPTG) overnight at 18°C. The cells were then harvested by centrifugation and resuspended in lysis buffer (50 mM Tris, pH 7.5, 500 mM NaCl, 10% [v/v] glycerol, 1 mM dithiothreitol [DTT] 0.01 mg/ml lysozyme, 0.006 mg/mL Dnase/RNase, protease inhibitor). The resuspended cells were disrupted by sonication and centrifuged at 14,000 × g for 45 min. Proteins were purified by a first step of Ni-NTA affinity chromatography with the elution buffer (50 mM Tris pH 7.5, 300 mM NaCl, 1 mM DTT, 250 mM imidazole). The eluted fractions were then concentrated and buffer exchange was performed in a centrifugal filter unit with gel filtration buffer (50 mM Tris pH7.5, 100 mM NaCl, and 1 mM DTT). The proteins were finally applied to a Superdex 200 16/600 GL size-exclusion column (GE Healthcare) and eluted with gel filtration buffer.
Crystallization, data collection, and structure determination
Request a detailed protocolCrystallization conditions were screened at 293 K using the sitting-drop vapor-diffusion method and commercial crystallization kits Crystal Screen 1 and 2, PEG/Ion 1 and 2 (Hampton Research). Further optimization screenings around conditions of initial hits were also performed. The crystallization drops (0.2 µL protein solution and 0.2 µL reservoir solution) were set up using a Mosquito crystallization robot and equilibrated against 70 µL reservoir solution. DltEextra and DltEextra-S128A were concentrated to around 10 mg/mL prior to crystallization. Diffraction quality crystals grew after around 1 wk in three different conditions: (1) 26% PEG MME 5K, 0.1 M Tris pH8.5, 0.15 M LiSO4 and 30% ethylene glycol (named hereafter apo condition); (2) 26% PEG 3350, 0.1 M ammonium tartrate (tartrate condition); and (3) 20% PEG 3350, 0.2 M ammonium tartrate, and 0.1 M TCEP HCl (TCEP condition). Crystals obtained in condition 2 were also soaked for 1 d in the crystallization condition containing 10 mM of LTA (prepared as described below) (LTA condition).
Crystals grown in conditions 2 and 3 were cryoprotected prior to data collection by rapid soaking in mother liquor containing 20% (v/v) glycerol and for the TA-soaked crystals 10 mM of TA. Crystals were flash-cooled in liquid nitrogen and diffraction data were collected at cryogenic temperature (100 K) on beamlines ID23-2 and ID30B at the European Synchrotron Radiation Facility (ESRF, Grenoble, France) and on beamlines PROXIMA-1 and PROXIMA-2 at SOLEIL synchrotron (Gif sur Yvette, France).
Data were processed using the XDS package (Kabsch, 2010). The space groups, asymmetric unit contents, and diffraction resolutions obtained for each of the crystallization conditions are presented in Supplementary file 1. The structures were solved by molecular replacement using Phaser implemented in PHENIX (Adams et al., 2010). The PDB entry 3WWX (Arima et al., 2016) was used as starting model to solve the DltEextra-apo structure derived from the crystals obtained in the crystallization condition 1. The other three structures (named DltEextra-tartrate; DltEextra-TCEP; and DltEextra-LTA) were solved using the DltEextra-apo as a starting model. Structures were refined using iterative rounds of COOT (Emsley et al., 2010) and PHENIX (Adams et al., 2010) or Refmac5 of CCP4 (Murshudov et al., 1997). The quality of the final structure was assessed with MOLPROBITY (Chen et al., 2010) before deposition at the Protein Data Bank under the accession codes 8AGR(DltEextra-apo), 8AIK(DltEextra-tartrate), 8AJI(DltEextra-TCEP), and 8AKH (DltEextra-LTA). Sequence alignments and structure images were generated with PyMOL (Schrödinger, LLC), UCSF ChimeraX (Pettersen et al., 2021), and ESPript and ENDscript (Robert and Gouet, 2014). Data collection and final refinement statistics are presented in Supplementary file 1.
Microscale thermophoresis assays
Request a detailed protocolProtein–ligand interactions were analyzed by MST (Jerabek-Willemsen et al., 2011). Buffer of purified and concentrated protein samples was exchanged on a desalting PD-10 column to labeling buffer containing HEPES 25 mM pH 7.5, NaCl 300 mM, Tween 20 0.05% (w/v). Proteins were then labeled with NHS red fluorescent dye according to the instructions of the RED-NHS Monolith NT Protein Labeling kit (NanoTemper Technologies GmbH, Munchen, Germany). After a short incubation of target-partner complex, the samples were loaded into MST premium glass capillaries and eight measurements were performed (four with target alone and four with target-partner complex) at 22°C. The assays were repeated three times for each binding check experiment. Data analyses were performed using Nanotemper Analysis software provided by the manufacturer.
Release of D-Ala from whole bacterial cells and quantification by UHPLC
Request a detailed protocolD-Ala esterified to teichoic acids was detected and quantified as described by Kovács et al., 2006. Briefly, D-Ala was released from lyophilized whole heat-inactivated bacteria by mild alkaline hydrolysis with 0.1 N NaOH for 1 hr at 37°C. After neutralization, the extract was incubated with Marfey’s reagent (1-fluoro-2,4-dinitrophenyl-5-L-alanine amide; Sigma). This reagent reacts with the optical isomers of amino acids to form diastereomeric N-aryl derivatives, which can be separated by HPLC. Separation of the amino acid derivatives was performed on a C18 reversed-phase column (Zorbax Eclipse Plus C18 RRHD 2.1 × 50 mm 1.8 µm Agilent) with an Agilent UHPLC 1290 system with a linear elution gradient of acetonitrile in 20 mM sodium acetate buffer pH 5.0. The eluted compounds were detected by UV absorbance at 340 nm. Quantification was achieved by comparison with D-alanine standards in the range of 50–2000 pmol. Mean values were obtained from three independent cultures with two injections for each.
Detection of LTA by western blot
Request a detailed protocolLTA detection was done as previously described (Webb et al., 2009) with some modifications. Bacterial cells from a 2 mL overnight culture in MRS were added to lysing matrix tubes containing 0.1 mm silica beads (Lysing Matrix B, MP Biomedicals) and were broken using an MP Biomedicals FastPrep 24 Homogenizer (4.5 m/s intensity for 30 s), with samples being kept on ice. After centrifugation of the suspension at 200 × g for 1 min to remove glass beads, cell envelopes and cell debris were recovered by centrifugation at 20.000 × g for 15 min. The pellet was resuspended in sample buffer containing 2% SDS. The sample buffer volume was normalized on the culture OD600nm. Samples were heated at 95°C for 20 min and centrifuged at 20.000 × g for 5 min. Supernatants were loaded on a 15% SDS-polyacrylamide electrophoresis (SDS-PAGE) gel. LTA were detected by western blotting after transfer onto a PVDF membrane (Immobilon-P transfer membrane, Millipore) by incubation with an anti-LTA mouse monoclonal antibody specific for poly-glycerolphosphate chains (clone 55, Origene, AM26274LE-N) at 1/1000 dilution, followed by anti-mouse antibody coupled to horseradish peroxidase (Thermo Fisher Scientific) at 1/2000. Western blot was revealed by chemiluminescence detection using Pierce ECL Western blotting substrate (Thermo Fisher Scientific) and a ChemiDoc imaging system (Bio-Rad).
Preparation of L. plantarum cell walls
Request a detailed protocolL. plantarum strains (500 mL) were grown overnight in MRS medium. Cell walls were prepared as described previously (Matos et al., 2017), with some modifications. pH of buffer solutions was kept ≤6.0 in the different steps to avoid potential D-Ala hydrolysis from teichoic acids. Briefly, bacteria inactivated by heat treatment were boiled in 5% SDS in 50 mM MES buffer pH5.5 for 25 min. After centrifugation for 10 min at 20.000 × g, the pellets were resuspended in 4% SDS in 50 mM MES buffer pH 5.5 and boiled again for 15 min. The pellet was washed six times with 10 mM MES pH 5.5 preheated at 60°C. An additional step was added consisting in shearing sacculi with glass beads. Pellets were resuspended in 1 mL of 10 mM MES pH 5.5, and the suspension was added to lysing matrix tubes containing 0.1 mm silica beads (MP Biomedicals). The cells were broken using an MP Biomedicals FastPrep 24 Homogenizer (4.5 m/s intensity for 30 s). After centrifugation of the suspension at 200 × g for 1 min to remove glass beads, insoluble material containing cell walls was recovered by centrifugation 20.000 × g for 15 min. The pellet was resuspended in 50 mM MES pH 6.0, further treated with pronase, trypsin, DNase, RNase, and lipase, and finally boiled in 4% SDS in 10 mM MES pH 5.5 for 15 min. The final pellet was washed four times with 10 mM MES pH 5.5 and twice with MilliQ H2O to remove SDS traces. The purified cell walls (containing PG, WTA, and polysaccharides) was lyophilized and further used for WTA and PG purification.
PG purification
Request a detailed protocolPurified cell walls were treated with hydrofluoric acid (48%) for 19 hr at 0°C to remove WTA and polysaccharides. The remaining insoluble purified PG was washed twice times with 250 mM Tris-HCl pH 8.0 and four times with MilliQ H2O and lyophilized.
WTA purification
Request a detailed protocolWTA were extracted from purified cell walls with TCA as described previously (Tomita et al., 2009) with some modifications. Briefly, lyophilized cell walls (50 mg; prepared as described above, keeping pH < 6.0 to avoid D-Ala hydrolysis from WTA) were incubated with 1 mL of 10% TCA at 4°C for 48 hr under rotation. The suspension was then centrifuged at 20.000 × g for 20 min at 4°C and the supernatant was recovered. WTAs were precipitated by addition of 5 V of ethanol and incubation overnight at –20°C. The pellet was purified by resuspension in TCA 10% and precipitation with ethanol. The final pellet was washed twice with cold ethanol, and the pellet was resuspended in MilliQ H2O and lyophilized. WTAs were resuspended in 100 mM ammonium acetate buffer pH 4.8 (buffer A) and purified by anion exchange chromatography on a 1 mL HiTrap Q HP column (Cytiva) equilibrated with buffer A. WTAs were eluted with a gradient from 0 to 100% buffer B (buffer A containing 1 M NaCl) in 20 min. Fractions were collected and the presence of WTA was assessed in microplates with thymol-H2SO4 reagent (Engelhardt and Ohs, 1987), allowing detection of hexoses substituents of WTAs. The WTA-containing fractions were pooled and dialyzed against 10 mM ammonium acetate pH 4.8 with Float-a-Lyzer G2 dialysis devices (cut-off 500–1000 Da) (Spectra/Por) and lyophilized.
LTA purification
Request a detailed protocolL. plantarum strains (5 L) were grown overnight in MRS. Bacteria were harvested at 4.500 × g for 10 min. The pellet was resuspended in 5 mL of 20 mM ammonium acetate buffer pH 4.6 (around 20 OD/mL). Bacteria were broken with a Constant Systems Ltd Basic Z Cell Disrupter (CellD) at 2000 bars. Non-broken cells were discarded by centrifugation at 3.000 × g for 15 min. LTA was obtained by butanol extraction allowing to keep D-Ala esterified on LTAs as described previously (Gründling and Schneewind, 2007; Morath et al., 2001). The supernatant was recovered and butanol-1 (1:1 V/V) was added in a glass container. The mixture was stirred for 30 min at room temperature. Insoluble material was discarded after centrifugation at 13.000 × g for 20 min in PPCO tubes (Nalgene). The liquid phase was recovered and centrifuged again; the aqueous phase (lower one) containing LTA was recovered. The sample was treated with DNAse II (Sigma D8764) at 50 µg/mL for 2 hr at 37°C to degrade contaminating DNA and lyophilized. LTA was further purified by hydrophobic interaction chromatography (HIC) with a Hi-prep Octyl FF 16/10 column with an AKTA chromatography system. LTA was dissolved in buffer A (10 mM ammonium acetate pH 4.7 containing 15% propanol-1). Elution was performed with a gradient of buffer B (10 mM ammonium acetate pH 4.7 containing 60% propanol-1) from 0 to 100% in 1 hr. Fractions were collected and the presence of LTA was tested by dot blot. Aliquots were spotted on a PVDF membrane (Immobilon-P transfer membrane, Millipore) and incubated with anti-LTA monoclonal antibody (Origene, AM26274LE-N) at 1/1000 dilution, followed by anti-mouse antibody coupled to horseradish peroxidase (Thermo Fisher) at 1/2000. Dot blot was revealed by chemiluminescence detection using Pierce ECL Western blotting substrate (Thermo Scientific) and detection with a Chemidoc system (Bio-Rad). Fractions containing LTA were pooled, dialyzed against 10 mM ammonium acetate pH 4.6 with Float-a-Lyzer G2 dialysis devices (cut-off 500–1000 Da) (Spectra/Por 500–1000 Da cut-off), and lyophilized.
PG, WTA, and LTA composition analysis and quantification
Request a detailed protocolPG was quantified by analysis of muramic acid (Mur) content after acid hydrolysis as described previously (Matos et al., 2017). PG (400 µg) was hydrolyzed by 6 N HCl for 16 hr at 100°C under vacuum. Mur was quantified by high-performance anion exchange chromatography coupled with pulse amperometric detection (HPAEC-PAD) with an ICS5000 system (Thermo Scientific) and a Dionex CarboPac PA-1 anion exchange column (4 × 250 mm) (Thermo Scientific) with a guard column. Quantification was made with a standard curve of pure Mur (Sigma-Aldrich) (50–1000 pmol).
WTA composition was obtained after hydrolysis with hydrofluoric acid 48% at 0°C for 48 hr. After HF evaporation, samples were further treated with 4 M trifluoroacetic acid (TFA) for 3 hr at 110°C. Composition was determined by HPAEC-PAD (ICS5000 system, Thermo Fisher Scientific) with a CarboPac PA20 column (Dionex). Composition and amount were determined by comparison with standard amounts of alditols (glycerol, ribitol) and monosaccharides glucosamine, galactosamine, glucose, galactose, rhamnose, and ribose.
LTA composition was obtained after hydrolysis by hydrofluoric acid 48% at 0°C for 48 hr. After HF evaporation, samples were treated with 4 M TFA for 3 hr at 110°C. Composition was determined by HPLC analysis with an Aminex HPX-87C column (Bio-Rad) with a Waters 2414 refractive index (RI) detector using alditols (glycerol, ribitol), monosaccharides and phosphate standards. Quantification was performed by comparison with standard amounts of the same compounds. D-Ala esterified to LTA was detected after alkaline hydrolysis using Marfey’s reagent derivatization, as described above for whole cells.
For supplementation experiments in fly tests, 1 µg of each purified component (PG, WTA, and LTA) was used. For comparison of components extracted from LpNC8 and ΔdltXABCD, the final PG, WTA, and LTA suspensions were adjusted to the same amount of Mur, Rbo, and Gro, respectively.
NMR analysis
Request a detailed protocolLTA was purified after butanol extraction allowing to keep D-Ala esterified on LTAs as described above. WTA was purified after TCA extraction from cell walls as described above. pH was kept below 6.0 at all purification steps of both polymers to avoid hydrolysis of the ester bond between D-Ala and the teichoic acid backbone chains. Samples were solubilized in highly enriched deuterated water (99.96% deuterium; EurisoTop, St-Aubin, France) and lyophilized. This process was repeated twice. Data were recorded on a 9.4 T spectrometer and a 18.8 T spectrometer were 1H resonated at 400.33 and 800.12 MHz, and 13C resonated at 100.2 and 200.3 MHz, respectively. All samples were inserted in 3 mm tubes with matching amounts of D2O. Acetone was added as an internal standard, starting from a solution of 2.5 μL of acetone–10 mL of D2O. All pulse sequences were taken from the Bruker library of pulse programs and then optimized for each sample. Spectral widths were 12 and 200 ppm for the proton and carbon observations, respectively. TOCSY was performed with various mixing times of 40–120 ms. Edited 1H-13C HSQC spectra were recorded with 1536 data points for detection and 256 data points for indirect direction.
Test of carboxyesterase activity on purified muropeptide
Request a detailed protocolDisaccharide-depsipentapeptide (GlcNAc-MurNAc-L-Ala-D-Gln-mDAP-L-Ala-L-Lac) was purified from PG of dacA1dacA2 double mutant of L. plantarumNC8. PG was digested with mutanolysin (Sigma-Aldrich), and the resulting soluble muropeptides were reduced by NaBH4 as described previously (Bernard et al., 2011). The reduced muropeptides were separated by reverse-phase ultra-high-pressure liquid chromatography (RP-UHPLC) with a 1290 chromatography system (Agilent Technologies) and a Zorbax RRHD Eclipse Plus C18 column (100 by 2.1 mm; particle size, 1.8 µm; Agilent Technologies) at 50°C using ammonium phosphate buffer and methanol linear gradient. The eluted muropeptides were detected by UV absorbance at 202 nm. The peak corresponding to the disaccharide-depsipentapeptide was collected. Purified muropeptide was then incubated with 10 µg of purified DltEextra or DacA1 in 50 mM Tris-HCl, 100 mM NaCl buffer overnight at 37°C. The reaction mixtures were analyzed by RP-UHPLC as described above and by LC-MS with an UHPLC instrument (Vanquish Flex, Thermo Scientific) connected to a Q Exactive Focus mass spectrometer (Thermo Scientific). Mass spectra were collected over the range m/z = 380–1400. Data were processed using Xcalibur QualBrowser v2.0 (Thermo Scientific).
Test of D-Ala esterase activity on LTA
Request a detailed protocolPurified DltEextra (100 and 200 µg), DltES128Aextra (100 µg), or DacA1 (100 µg) were incubated overnight at 37°C with 200 µg D-Ala-LTA purified from L. plantarumNC8 in a final volume of 100 µL of 20 mM MES, 150 mM NaCl pH 6.0. As a control, 200 µg of D-Ala-LTA was incubated in the same conditions without enzyme. After lyophilization, samples were treated with 48% hydrofluoric acid for 48 hr at 0°C, allowing LTA depolymerization without release of D-Ala esterified on Gro residues. Gro-D-Ala subunits and free D-Ala released from LTA chains were analyzed by LC-MS using an UHPLC instrument (Vanquish Flex, Thermo Scientific) connected to a Q Exactive Focus mass spectrometer (Thermo Fisher Scientific). A reverse-phase column (Hypersyl Gold AQ C18 column; 200 by 2.1 mm; particle size 1.9 µm; Thermo Fisher Scientific) was used for separation. To enhance the retention and resolution of the column, heptafluorobutyric acid (HFBA) was used as an ion-pairing agent. Samples were diluted 20-fold in buffer A with 2 µL injected onto the column. Buffer A contained 0.2% HBFA in MilliQ H2O and buffer B contained 0.2% HBFA in acetonitrile/MilliQ H2O (80:20, V/V). Elution was performed at flow rate 0.3 mL/min using an isocratic step of buffer A for 3 min followed by a gradient to 5% of buffer B in 5 min. Mass analysis was performed in single ion monitoring (SIM) mode by following Ala and Ala-Gro with the respective m/z of 90.05 and 164.09. Ala and Gro-Ala were identified by their MS and MS2 spectra.
Data availability
Diffraction data have been deposited in PDB under the accession codes 8AGR/8AIK/8AJI/8AKH-All data generated during this study are included in the manuscript and supporting file and a single Source Data file has been provided.
-
RCSB Protein Data BankID 8AGR. Crystal structure of DltE from L.plantarum, apo form.
-
RCSB Protein Data BankID 8AIK. Crystal structure of DltE from L.plantarum, tartare bound form.
-
RCSB Protein Data BankID 8AJI. Crystal structure of DltE from L.plantarum, TCEP bound form.
-
RCSB Protein Data BankID 8AKH. Crystal structure of DltE from L.plantarum soaked with LTA.
References
-
PHENIX: a comprehensive python-based system for macromolecular structure solutionActa Crystallographica Section D Biological Crystallography 66:213–221.https://doi.org/10.1107/S0907444909052925
-
Gut microbiome development and childhood undernutritionCell Host & Microbe 30:617–626.https://doi.org/10.1016/j.chom.2022.04.002
-
Characterization of O-acetylation of N-acetylglucosamine: a novel structural variation of bacterial peptidoglycanThe Journal of Biological Chemistry 286:23950–23958.https://doi.org/10.1074/jbc.M111.241414
-
molprobity: all-atom structure validation for macromolecular crystallographyActa Crystallographica Section D Biological Crystallography 66:12–21.https://doi.org/10.1107/S0907444909042073
-
Features and development of cootActa Crystallographica. Section D, Biological Crystallography 66:486–501.https://doi.org/10.1107/S0907444910007493
-
Trace analysis of sugars by HPLC and post-column derivatizationChromatographia 23:657–662.https://doi.org/10.1007/BF02311495
-
How commensal microbes shape the physiology of Drosophila melanogasterCurrent Opinion in Insect Science 41:92–99.https://doi.org/10.1016/j.cois.2020.08.002
-
Molecular interaction studies using microscale thermophoresisASSAY and Drug Development Technologies 9:342–353.https://doi.org/10.1089/adt.2011.0380
-
Integration, scaling, space-group assignment and post-refinementActa Crystallographica Section D Biological Crystallography 66:133–144.https://doi.org/10.1107/S0907444909047374
-
The digestive tract of Drosophila melanogasterAnnual Review of Genetics 47:377–404.https://doi.org/10.1146/annurev-genet-111212-133343
-
Efficient insertional mutagenesis in lactococci and other gram-positive bacteriaJournal of Bacteriology 178:931–935.https://doi.org/10.1128/jb.178.3.931-935.1996
-
Structures of two kinetic intermediates reveal species specificity of penicillin-binding proteinsJournal of Molecular Biology 322:111–122.https://doi.org/10.1016/S0022-2836(02)00742-8
-
Structure–function relationship of cytokine induction by lipoteichoic acid from Staphylococcus aureusJournal of Experimental Medicine 193:393–398.https://doi.org/10.1084/jem.193.3.393
-
Crystal structure of a peptidoglycan synthesis regulatory factor (PBP3) from Streptococcus pneumoniaeThe Journal of Biological Chemistry 280:15984–15991.https://doi.org/10.1074/jbc.M408446200
-
Refinement of macromolecular structures by the maximum-likelihood methodActa Crystallographica Section D Biological Crystallography 53:240–255.https://doi.org/10.1107/S0907444996012255
-
Stunting of growth in developing countriesNestle Nutr Works Se 125:14–27.https://doi.org/10.1159/isbn.978-3-318-07043-9
-
A continuum of anionic charge: structures and functions of D -alanyl-teichoic acids in Gram-positive bacteriaMicrobiology and Molecular Biology Reviews 67:686–723.https://doi.org/10.1128/MMBR.67.4.686-723.2003
-
Lipoteichoic acid synthesis and function in gram-positive bacteriaAnnual Review of Microbiology 68:81–100.https://doi.org/10.1146/annurev-micro-091213-112949
-
Incorporation of D-alanine into lipoteichoic acid and wall teichoic acid in Bacillus subtilis: identification of genes and regulationThe Journal of Biological Chemistry 270:15598–15606.https://doi.org/10.1074/jbc.270.26.15598
-
Deciphering key features in protein structures with the new endscript serverNucleic Acids Research 42:W320–W324.https://doi.org/10.1093/nar/gku316
-
The gram-positive bacterial cell wallMicrobiology Spectrum 7:2018.https://doi.org/10.1128/microbiolspec.GPP3-0044-2018
-
Structural characterization of teichoic acids from Lactobacillus brevisCarbohydrate Research 345:538–542.https://doi.org/10.1016/j.carres.2009.12.007
-
Nih image to imagej: 25 years of image analysisNature Methods 9:671–675.https://doi.org/10.1038/nmeth.2089
-
Gut microbiota and host juvenile growthCalcified Tissue International 102:387–405.https://doi.org/10.1007/s00223-017-0368-y
-
Structures of two monomeric units of teichoic acid prepared from the cell wall of Lactobacillus plantarum NRIC 1068Bioscience, Biotechnology, and Biochemistry 73:530–535.https://doi.org/10.1271/bbb.80582
Article and author information
Author details
Marie-Pierre Chapot-Chartier

Funding
Agence Nationale de la Recherche (ANR-18-CE15-0011)
- François Leulier
- Marie-Pierre Chapot-Chartier
- Christophe Grangeasse
Fondation pour la Recherche Médicale (DEQ20180839196)
- François Leulier
Fondation Innovations en Infectiologie
- Renata C Matos
ENS de Lyon (Emergence)
- Renata C Matos
Fondation Bettencourt Schueller
- Christophe Grangeasse
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
The authors thank Octobre Clocher, Dali Ma, and Cathy Ramos for their contribution to the early stages of the bacteriology and Drosophila tasks of this project, Céline Freton for help in microscale thermophoresis, Laurane Bernelin for her contribution at the initial stages of the DltE production and crystallization, and Emmanuel Maes and Jessica L Davis for technical assistance. We acknowledge the SFR Biosciences (UAR3444/US8) for access to the Protein Science (crystallogenesis robots) and the ArthroTools platform (Drosophila facility) and support on the beamlines ID23-2 and ID30B at the European Synchrotron Radiation Facility (ESRF, Grenoble, France) and on PROXIMA-1 and PROXIMA-2A at SOLEIL synchrotron (Gif sur Yvette, France), the PAGes core facility (http://plateforme-pages.univ-lille1.fr) and UAR 2014 /US 41 (Plateformes Lilloises en Biologie & Santé) for providing the scientific and technical environment conducive to achieve this work. We acknowledge ChemSyBio (Micalis, INRAE, France) for access to mass spectrometry facilities. This work was funded by the collaborative grant ANR-18-CE15-0011 to FL, MPCC and CG. Work in the FL lab was also supported by a FRM grant (DEQ20180839196), a FINOVI starting grant to RCM and an ENSL Emergence grant to RCM. Work in the CG lab was also supported by the Fondation Bettencourt-Schueller.
Copyright
© 2023, Nikolopoulos, Matos, Ravaud et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,459
- views
-
- 266
- downloads
-
- 14
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 14
- citations for umbrella DOI https://doi.org/10.7554/eLife.84669
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Structure–function analysis of Lactiplantibacillus plantarum DltE reveals D-alanylated lipoteichoic acids as direct cues supporting Drosophila juvenile growth
eLife 12:e84669.
https://doi.org/10.7554/eLife.84669




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}