A phenotypic screening platform utilising human spermatozoa identifies compounds with contraceptive activity
- University of Dundee, United Kingdom
- Ninewells Hospital and Medical School, University of Dundee, United Kingdom
Abstract
There is an urgent need to develop new methods for male contraception, however a major barrier to drug discovery has been the lack of validated targets and the absence of an effective high-throughput phenotypic screening system. To address this deficit, we developed a fully-automated robotic screening platform that provided quantitative evaluation of compound activity against two key attributes of human sperm function: motility and acrosome reaction. In order to accelerate contraceptive development, we screened the comprehensive collection of 12,000 molecules that make up the ReFRAME repurposing library, comprising nearly all the small molecules that have been approved or have undergone clinical development, or have significant preclinical profiling. We identified several compounds that potently inhibit motility representing either novel drug candidates or routes to target identification. This platform will now allow for major drug discovery programmes that address the critical gap in the contraceptive portfolio as well as uncover novel human sperm biology.
Introduction
An effective and comprehensive family planning strategy is fundamental to delivery on the United Nations 17 Sustainable Development Goals (Starbird et al., 2016; Goodkind et al., 2018), however the current contraceptive portfolio is suboptimal. For example, it is estimated that more than 214 million women in developing countries have an unmet need for contraception which results in 89 million unintended pregnancies and 48 million abortions every year, with substantial impacts on maternal and child health and poverty levels (Guttmacher Institute, 2017). To achieve the UN targets it is critical that involvement of men be considered of equal importance as women (Hardee et al., 2016). However, it is noticeable that no effective, reversible and widely available form of contraception has been developed for men since the condom and as such the burden falls largely on the woman. The development of drugs that can be used in the male would address a critical gap in the contraceptive portfolio.
Progress in developing new male contraceptives has been slow for a plethora of reasons. Despite decades of research, safe and effective hormonal approaches to prevent spermatozoa production have yet to be realised and viable alternatives remain elusive (Page and Amory, 2018). Developing a drug for any purpose is difficult but one that blocks sperm function is a particular challenge as spermatozoa don’t divide, transcribe or translate, have specialized organelles, are highly motile and continually produced in large numbers (~100 million per day). Moreover, as the production of fertilization competent cells is necessarily different between species, it is essential to use a human system early in the drug discovery cascade to increase the chances of translational success. The spermatozoa’s sole function post-ejaculation is to swim until it closes in on the oocyte. The spermatozoa are then able to undergo a unique exocytotic process called the acrosome reaction (AR) which facilitates penetration of the zona pellucida, the oocyte’s outer glycoprotein shell and allows fusion with the oocyte’s plasma membrane, delivering its DNA. Hence, a male contraceptive could work in a number of different ways – it could completely halt spermatogenesis, block an essential function of the spermatozoon (such a motility) or perturb a cellular process that results in a non-productive interaction with the oocyte (such as prematurely triggering the acrosome reaction, and/or by blocking exocytosis or inhibiting receptor binding). Low sperm motility is a common cause of male infertility (Samplaski et al., 2019) indicating that targeting this process should yield effective drugs. The options for delivery of a contraceptive are wide ranging, but our primary aim is to develop a relatively fast-acting safe drug, self-administered by the male, that specifically and effectively irreversibly blocks the sperm’s ability to get to the egg and/or fertilise it.
Though some potential sperm-specific contraceptive drug targets, such as the calcium channel CatSper, have been identified (Lishko and Mannowetz, 2018), our relatively poor understanding of spermatozoon biology makes target-agnostic, or phenotypic strategies more attractive (Barratt et al., 2017). Phenotypic drug discovery is undergoing a renaissance in a number of therapeutic areas because it can uncover novel biology in an unbiased way. Retrospective analysis of drug approvals shows the approach to be more successful in first-in-class medicine discovery than previously appreciated (Swinney and Anthony, 2011; Moffat et al., 2017). However, for male contraceptive drug discovery, a limiting barrier has been the absence of a scalable drug screening system using spermatozoa.
To address this unmet need we set out to develop the first high-throughput phenotypic screening platform for sperm where an imaging-based kinetic analysis module that tracks motility, is followed by a flow cytometry -based assay module that detects both the degree of acrosome reaction (AR) and cell viability - both modules coordinated by a fully automated robotic system. The bespoke computational pipeline quantitatively evaluates each compound’s effect on both motility and AR (and viability) thus achieving unprecedented throughput. The platform was used to identify motility inhibitors from a unique 12,000 molecule ReFRAME (Repurposing, Focused Rescue, and Accelerated Medchem) library, which represents the most comprehensive collection of drugs available, as it contains nearly all the small molecules that have achieved regulatory approval and others that have undergone varying degrees of clinical development or have had significant preclinical profiling (Janes et al., 2018). This advance opens up the possibility of accelerated male contraceptive development by allowing drug repurposing, target identification as well as screening of chemical diversity libraries for novel medicinal chemistry start points.
Results
Phenotypic assay development
Our automated platform (Figure 1A) allows for the identification of relatively fast-acting compounds with a rapid plate-processing time (~75 minutes/ 384-well plate), where the motility assay runs for ~30 min followed directly by the AR assay (~45 min). Automated cell tracking (Figure 1B and Figure 1—figure supplement 1A,B) using time-lapse images allowed kinetic parameter quantification on >200 cells/well (Figure 1C). The kinetics show good agreement with clinical systems (Figure 1—figure supplement 1C) and a comparable distribution was observed between experimental days, plates and positions (Figure 1—figure supplement 2) with any small differences between days stemming from donor pool variance. Importantly, spermatozoa were found to be tolerant to the DMSO concentrations used in our screening program (0.0625%–0.1% Figure 1—figure supplement 1D). To determine the optimal screening batch size untreated spermatozoa were dispensed every 30 min for 150 min observing only a small decrease in motility (~10%) after four plates were screened (Figure 1—figure supplement 1E). A flow cytometry-based assay measuring AR (Figure 1D) was run directly afterwards (Figure 1E and Figure 1—figure supplement 1F).
Figure 1 with 2 supplements see all
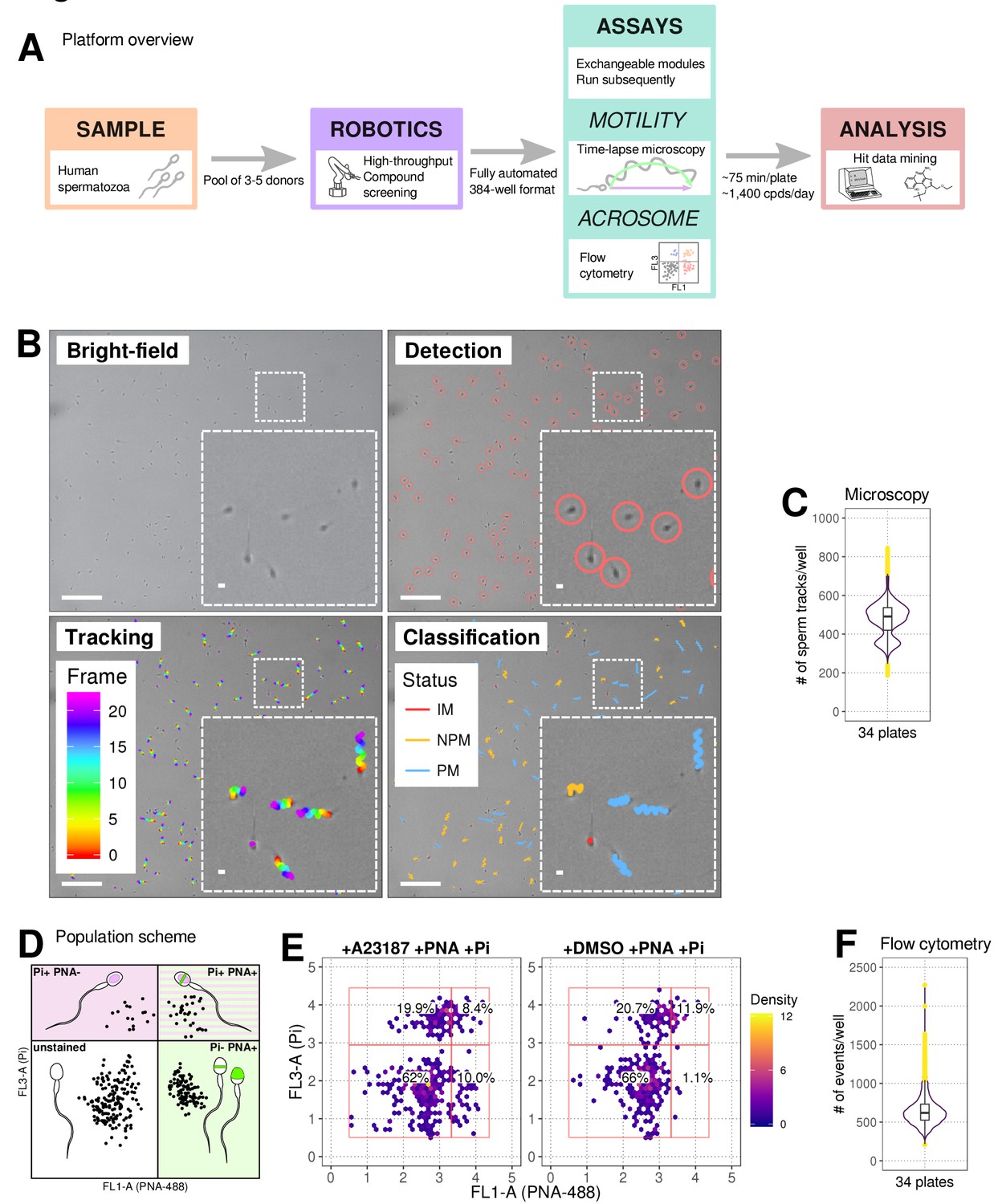
Phenotypic Assay workflows.
(A) Graphical summary of modular screening workflow where motility measurement is followed by acrosome reaction (AR) measurement allowing a screening throughput of >1400 compounds per donor pool (B) Steps in imaging and analysis: human sperm are recorded with brightfield illumination (first panel) then sperm heads detected using a particle tracking algorithm (second panel) which are then tracked across the timelapse series of images (third panel) and subsequently classified (fourth panel). Each panel contains a zoomed-in subsection of the field. Colour coding for tracking distance and kinetic classification is shown in the panel insets: Tracking panel - rainbow gradient (showing progression over time); Classification panel - red for immotile (IM), yellow for non-progressively motile (NPM) and blue for progressively motile (PM). Scale bars: 100 μm (main images), 5 μm (insets). (C) Sperm counts per well after microscopy and detection shown in a combined violin/box plot. Colours: purple violin outline (probability density of values), yellow dots (outliers of boxplot). (D) Graphical summary of the expected populations determined by flow cytometry based on distribution of cells measured with FL3-A (Pi) vertical axis and FL1-A (PNA-488) horizontal axis: dead cells (upper left, Pi+ PNA-); dead and acrosome-reacted (upper right, Pi+ PNA+); unstained/live/non-reacted (lower left) and live acrosome-reacted (lower right, Pi-PNA+). (E) Example flow cytometry data comparing sperm treated with the Ca2+ ionophore (A23187) which induces AR (left panel) with sperm from DMSO-treated well (right panel). Colours indicate event density. (F) Combined violin/box plot data showing flow cytometry event counts per well. Colours and label as in (C).
Screening of the ReFRAME library, confirmation of hits by dose response
The ReFRAME library (11,968 compounds) was supplied in ‘assay-ready’ imaging plates and was solubilised prior to addition of sperm. High assay quality was achieved throughout with a Z’-factor between 0.4–0.8 (Figure 2B). A total of 63 putative hit compounds decreased motility to varying degrees based on a hit selection cut-off of >15% reduction in motility (Figure 2A with examples in Figure 2C,D,F,G and Video 1 and 2). 14 compounds were selected as putative AR+ hits (Figure 3A with examples in B, C). Motility hits were confirmed with resupplied material in dose-response experiments. There was a dose-dependent decrease in motility for 29 compounds (46% hit confirmation rate), with EC50 values as low as 0.05 µM and effect sizes ranging from 15–100% (Figure 2F,G; Figure 2—source data 1, Supplementary file 1). Amongst the confirmed hits were the aldehyde dehydrogenase inhibitor, Disulfiram (70% maximum reduction at 10 µM) and a putative platelet aggregation inhibitor, KF-4939 (100% max. reduction; EC50 = 0.49 μM), and a range of other compounds having very modest effect and/or showing low potency (See Figure 2—source data 1, Supplementary file 1). In order to test reversibility, wash out experiments were performed. Only a small recovery of motility was observed, after wash out of Disulfiram (Figure 2—figure supplement 1). In the AR screen, 9 of the resupplied compounds had a dose-dependent effect with EC50 values as low as 0.4 µM (See Figure 3—source data 2, Supplementary file 2). However, following the orthogonal assay triaging we implemented to eliminate assay interference compounds, none of the AR hits were found to be true positives (see examples in Figure 3—figure supplement 1D) despite the primary screen showing good assay robustness.
Figure 2 with 1 supplement see all
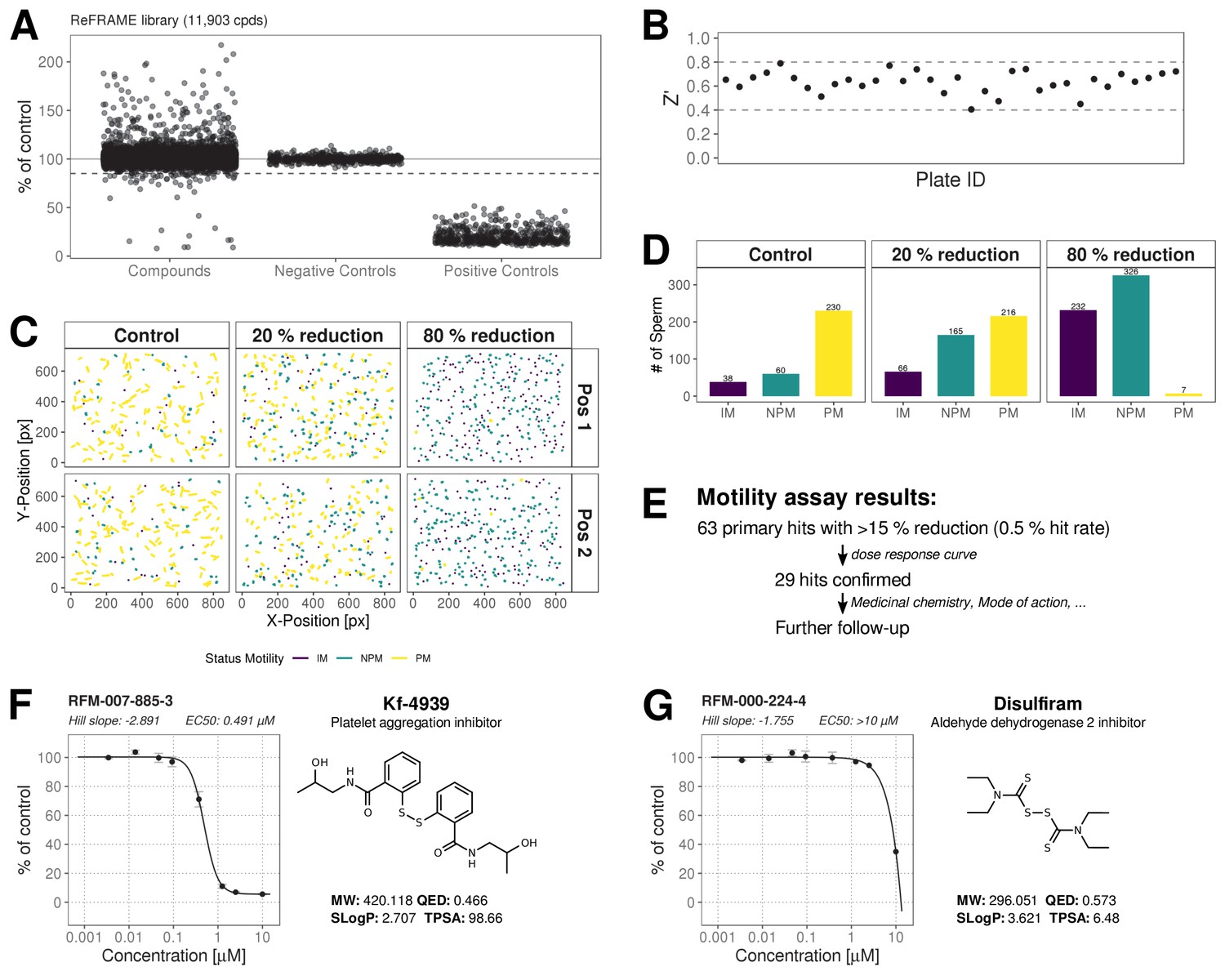
ReFRAME Library Screening: Motility Assay Results.
(A) Primary screening results of the motility assay. Each dot represents a well (either compound or control well) showing % of DMSO control (Curvilinear Velocity: VCL). Positive controls (Pristimerin), negative controls (DMSO) and individual compound datapoints are shown. The dashed line showing the 15% of control (reduction in VCL) – the cut-off for primary hit selection. Total number of compounds = 11,903, excluding wells with auto-focus errors and with ‘sticky’ compounds which have been excluded from analysis. See dataset (http://doi.org/10.5061/dryad.jdfn2z36z) Figure 2—source code 1. (B) Assay robustness: the standard high throughput screening metric, Z' (see Materials and methods; Zhang et al., 1999) was used to determine the performance of the assay for all screening plates. Dashed lines indicate min/max Z' values. (C) Tracking data visualizations of 3 example wells showing sperm tracks of both imaging positions (Position 1 [Pos. 1] and Position 2 [Pos. 2] respectively) within the wells. A DMSO control well (left panels ‘Control’) shows a large number of progressively motile (PM) sperm (yellow) with few non-progressively motile (NPM) sperm (green and very few immotile (IM) sperm (purple) – this is in contrast to the shorter tracks and higher levels of NPM and IM in the middle panel (‘20% reduction’) for a compound that shows 20% inhibition of motility (i.e. 80% of control) and the right hand panel (‘80% reduction’) for a compound showing 80% inhibition (i.e. 20% of control), showing almost all cells are in the IM and NPM classes. (D) Histogram of sperm tracks quantification of the data show in (C). (E) Summary of motility assay hit rate (0.5%) and reconfirmation rate (0.24%). (F-G) Dose response confirmation of two hits. 8-point 3-fold dilution curves are shown with 10 μM as the highest concentration. Two data points per concentration (n = 2, data point is mean ± SD). Each curve is a 4-parameter logistic fit. Each plot shows estimated values Hill Slope and EC50. The chemical structure of the hit compound is shown as well as some annotation and physicochemical properties. Physicochemical properties were calculated using RDKit, Python and KNIME: SlogP = partition coefficient (Wildman and Crippen, 1999); TPSA is the Topological Polar Surface Area (Ertl et al., 2000); MW is the exact Molecular weight; QED = Quantitative Estimate of Drug-likeness (Bickerton et al., 2012).
-
Figure 2—source code 1
R Code for Figure 2 primary motility assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig2-code1-v2.r
-
Figure 2—source code 2
R Code for Figure 2—source data 1 dose response confirmation motility assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig2-code2-v2.r
-
Figure 2—source data 1
Dose response and additional data for primary motility hits.
Compound names and nominal target/functional annotation shown on left along with chemical structure and physico-chemical properties. The 8-point dose response curves for each hit with estimated Hill slope, EC50 and Efficacy [% max reduction] are shown on the right. Two data points per concentration (n = 2); data points are Mean ± SD. A 4-parameter logistic fit has been performed using R package: dr4pl. Calculated EC50 values and Efficacy (% max effect) in the motility assay are shown in the right hand section. CC50 values for HEK293 and HepG2 CellTiter-Glo cytotoxicity assays performed by CALIBR (www.reframedb.org) are shown in the right hand section. Note: 0 = inactive in cytotoxicity assay. The chemical structure of the hit compound is shown as well as some annotation and physicochemical properties (for nomenclature description see Figure 2). See Figure 2—source data 2 along with Figure 2—source code 2.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig2-data1-v2.pdf
-
Figure 2—source data 2
Dose response confirmation data motility assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig2-data2-v2.csv
Figure 3 with 1 supplement see all
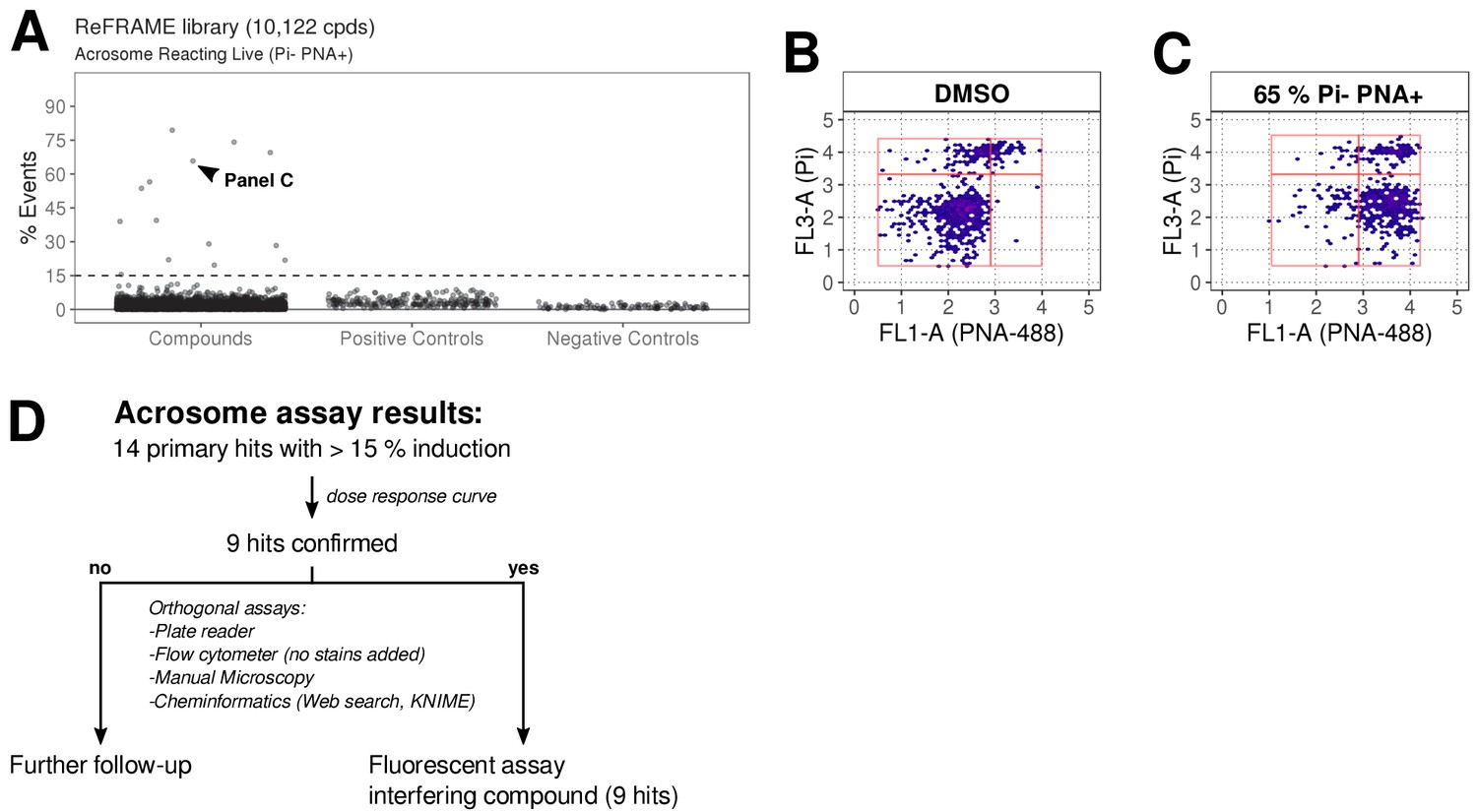
ReFRAME library screening: ar assay results.
(A) Results of primary screening of the library using the acrosome assay (live cells, acrosome reacting, Pi- PNA+ population). Each dot represents a well (either compound or control well). Shown is % Events (number of events in Pi- PNA+ gate relative to total events in the sampled well). Datapoints for compounds, negative controls (DMSO) and positive controls (A23187) are shown. Black dashed line = 15% threshold for primary hits selection. See Figure 3—source data 1 along with Figure 3—source code 1. (B-C) Data from two example wells: a DMSO well (left panel) and a well with 65% Pi- PNA+ population (right panel). (D) Summary of acrosome assay results before and after triage.
-
Figure 3—source code 1
R Code for Figure 2 primary acrosome assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig3-code1-v2.r
-
Figure 3—source code 2
R Code for Figure 3—source data 2 dose response confirmation acrosome assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig3-code2-v2.r
-
Figure 3—source data 1
Primary screening data acrosome assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig3-data1-v2.csv
-
Figure 3—source data 2
Dose response and additional data for primary AR hits.
Compound names and nominal target/functional annotation shown on left along with chemical structure and physico-chemical properties. The 8-point dose response curves for each hit with estimated Hill slope, EC50 and Efficacy [% max induction] values are shown on the right. Two data points per concentration (n = 2); data points are Mean ± SD. A 4-parameter logistic fit has been performed using R package: dr4pl. CC50 values for HEK293 and HepG2 CellTiter-Glo cytotoxicity assays performed by CALIBR (www.reframedb.org) are shown in the right hand section. Note: 0 = inactive in cytotoxicity assay. Physicochemical properties were calculated using RDKit, Python and KNIME: SlogP = partition coefficient (Wildman and Crippen, 1999); TPSA is the Topological Polar Surface Area (Ertl et al., 2000); MW is the exact Molecular weight; QED = Quantitative Estimate of Drug-likeness (Bickerton et al., 2012). See Figure 3—source data 3 along with Figure 3—source code 2 .
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig3-data2-v2.pdf
-
Figure 3—source data 3
Dose response confirmation data acrosome assay.
- https://cdn.elifesciences.org/articles/51739/elife-51739-fig3-data3-v2.csv
Video 1
Movie was generated using a brightfield image sequence for a typical control well (left pane), a well where a compound reduced motility by 20% (middle pane) and one where it reduced it by 80% (right pane).
Video 2
Movie was generated using a brightfield image sequence for a typical control well (left pane), a well where a compound reduced motility by 20% (middle pane) and one where it reduced it by 80% (right pane).
Tracking is overlayed in each panel. Colour coding is detailed in Figure 2.
Discussion
A major barrier in the search for new male contraceptives has been the lack of an effective high throughput phenotypic screening system. This study presents the first platform based on imaging and flow cytometry that measures two fundamental aspects of sperm behaviour – motility and AR, allowing for the rapid screening of compound collections.
A significant challenge was to create a system for the tracking of these highly motile cells that was fully automated and scalable to allow throughput at sufficient speed. Moreover, concomitant assessment of a second functional attribute, AR along with cell viability was developed in order to maximise use of the biological material. Assessment of motility required novel implementation of tracking algorithms. Importantly, the kinetic outputs were equivalent to those from the ‘industry-standard’ CASA, which is very low throughput and unsuitable for screening more than a few compounds at one time. Significant workflow optimisation resulted in good assay robustness and an acceptable hit confirmation rate following the primary screen using a medium stringency cut off for hit selection (>15% reduction). For AR detection, a flow cytometry approach was taken based on established methods (Mortimer et al., 1987) and showed good assay robustness. However, the presence of assay-interfering compounds in screening libraries (potential fluorescent compounds including DNA intercalators) necessitated further triaging steps (see Materials and methods) to eliminate false positives. This combination of triaging approaches meant that, in this screen, no AR compounds were suitable for progression. An improvement to the AR primary assay could be to use a far-red labelled lectin reagent to detect the glycoproteins. Other variants of the assay would be to either use sperm that had been capacitated, a state of increased readiness to undergo the AR process or use it to find drugs that would block the AR induced by a physiological stimulus. These latter variants are worthy of future investigation.
It is worth noting that whilst the primary goal of this programme is to identify compounds that will act as contraceptive by decreasing motility we detect a larger number of putative motility upregulating hits. These compounds are of interest for not only target identification and uncovering novel sperm biology but also their potential to improve fertility treatments. Overall, although the two assays presented here are fairly complex biological assay using primary cells, it achieved acceptable throughput with the potential to increase batch size for larger screening campaigns.
The advantage of using the ReFRAME collection was the potential to repurpose drugs that are already approved for other indications, or are at earlier stages of clinical progression. The associated compound annotations and safety data offers the potential to help accelerate the development of an effective male contraceptive. In the motility assay the hit rate of ~0.2% is typical for a cell-based assay, however surprisingly a relatively high number of hits were not deemed suitable for compound progression. Examples included potentially toxic mercury-containing compounds (phenylmercuric borate and mercufenol chloride); antiseptics and antibiotics including tyrothricin, or compounds with impacts on fundamental biology for example the microtubule stabiliser and chemotherapeutic, docetaxel. However, several hits are potentially of interest. One is Disulfiram (70% reduction at 10 μM) a drug that is well tolerated, has long been used for treating alcohol dependency and has previously been shown to inhibit sperm motility (Lal et al., 2016). Disulfiram is unlikely to be developable as a suitable contraceptive drug due to its side-effects when alcohol is consumed but has potential to facilitate mode-of-action studies. One recent report identified 3-Phosphoglycerate dehydrogenase (PHGDH) as a target for disulfiram (Spillier et al., 2019) whilst another identified NLP4, a subunit of VCP/p97 segregase (Skrott et al., 2017; Skrott et al., 2019) although the relevance of either to sperm function is unclear. The identification of KF-4939, a putative platelet aggregation factor (PAF) inhibitor, as a potent hit (100% max. reduction EC50 = 0.5 µM) may also be of interest since PAF has been used to improve sperm function in male infertility treatments (Roudebush et al., 2004). It is noteworthy that like disulfiram, KF-4939 also contains a disulphide linkage indicating that its biological effect maybe mediated by modifying sulfhydryls on proteins (Yamada et al., 1985). Further thiol-containing compounds, bismuth ethanedithiol (98% max reduction; EC50 = 2.5 μM) and the anti-fungal benzothiazole, Ticlatone (96% max. reduction EC50 = 4.4 µM) were also potent hits, suggesting free cysteines in, as yet, unidentified protein(s), may play a critical role in motility. Of interest is the observation that the Toll-like receptor 7/8 ligand (Resquimod; R848), recently shown to preferentially reduce the motility of X chromosome-bearing mouse sperm by suppressing ATP production (Umehara et al., 2019), was a hit, demonstrating that this phenotypic screening approach has the potential to uncover new aspects of sperm biology.
It was somewhat surprising that relatively few drugs or drug-like molecules in the ReFRAME collection were potent and effective inhibitors in the motility assay. This may reflect the unique biology of the spermatozoon that has evolved for a singular purpose – to reach the oocyte and fertilise it and/or that the targets within pharma’s therapeutic areas of interest over the last few decades are not similarly functional in human sperm. Conversely however, we did observe a large number of compounds (Figure 2A) that had a significant positive effect on sperm motility which may be target agonists as well as potential start points for treating men with low sperm motility. The development of an effective safe male contraceptive is challenging because a prerequisite, other than the obvious need for safety, is a level of reversibility. Other possibilities, such as a drug taken by the female or an agent for use as a topical spermicide (for example to replace nonoxynol-9) are options to consider in the future and depend to a large extent on the pharmacokinetic properties of compounds we identify and their mode of action – for example some may have poor bioavailability.
In summary, our high-throughput phenotypic platform allows for the screening of bioactives including natural products, focussed small molecule libraries and chemical diversity sets, which can provide novel start-points for male contraceptive drug development, as well as the identification of new drug targets in human spermatozoa.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Biological sample (Homo sapien) | Live spermatozoa | Donated semen samples | Local ethical approval (13/ES/0091) | |
| Antibody | Lectin PNA (Arachis hypogaea), Alexa Fluor 448 Conjugate | ThermoFisher Scientific | ThermoFisher:L21409; RRID: AB_2315178 | Stored as 1 mg/mL stock in DMSO; used at 1:1000 final dilution |
| Commercial assay or kit | Propidium Iodide; Live/Dead Sperm Viability kit, | ThermoFisher Scientific | ThermoFisher:L7011, | Stored as 2.4 mM stock; used at a 1:2000 final dilution |
| Chemical compound, drug | ReFRAME (Repurposing, Focused Rescue, and Accelerated Medchem) library | CALIBR at the Scripps Institute; Publication (Janes et al., 2018) | www.reframedb.org | |
| Chemical compound, drug | Pristimerin | Merck | Merck:530070 | Stored as 10 mM stock in DMSO; used at final concentration of 20 µM |
| Chemical compound, drug | calcium ionophore (A23187) | Sigma-Aldrich | Sigma-Aldrich:C7522 | Stored as 10 mM stock in DMSO, used at a final concentration of 10 µM) |
| Chemical compound, drug | Disulfiram | Tocris | Tocris:3807 | Stored as 10 mM stock in DMSO, used at 10 μM final concentration |
| Software, algorithm | Trackpy v0.4.1 | Zenodo. (http://doi.org/10.5281/zenodo.1226458) | Publication: (Crocker and Grier, 1996); Publication: (Allan, 2018) | |
| Software, algorithm | FFMPEG | FFmpeg Developers (http://ffmpeg.org) | RRID:SCR_016075 | |
| Software, algorithm | Bioconductor packages | Bioconductor (https://bioconductor.org) | RRID:SCR_006442 | flowCore, flowDensity, flowWorkspace, ggcyto |
| Software, algorithm | HDF5 | HDF Group (www.hdfgroup.org) | ||
| Software, algorithm | dr4pl | Dr4pl (https://cran.r-project.org/web/packages/dr4pl/index.html) | ||
| Software, algorithm | Code used for data analysis | This paper | The R code used for data analysis is included in the supplement files accompanying this paper | |
| Software, algorithm | KNIME | Berthold et al., 2008 | https://www.knime.com | |
| Software, algorithm | RDKit | RDKit, 2018 | https://www.rdkit.org |
Ethical approval
Request a detailed protocolWritten consent was obtained from each donor in accordance with the Human Fertilization and Embryology Authority (HFEA) Code of Practice (version 8) under local ethical approval (13/ES/0091) from the Tayside Committee of Medical Research Ethics B.
Development of methods for motility and Acrosome Reaction (AR)
Request a detailed protocolA pre-existing automated cell-based phenotypic screening platform was adapted with the use of live human spermatozoa. The platform utilises a Yokogawa CV7000 Cell Voyager high-throughput microscope able to image 384 multiwell plates under full environmental control acquiring timelapse images at a sufficiently high speed (~48 fps)_to capture the fast-moving sperm for tracking analysis. For high throughput screening with human spermatozoa to be effective and yield meaningful hits, the system needed to replicate, as close as possible, the manual workflow currently used in an andrology lab, so that the effect of large numbers of compounds on viable human sperm could be performed. In order to achieve this goal, we investigated a number of factors that could impact the quality of the data such as: plate type; concentration and number of spermatozoa per well; logistics of spermatozoa preparation by density gradient centrifugation and handling; donor-pooling; dispense speed for getting spermatozoa into 384 well plates; timing of compound addition; temperature control; image acquisition parameters; algorithm choice for motility assessment and data management and, evaluating the overall speed of data analysis. In addition, a flow cytometry-based assay and data analysis workflow was developed to measure acrosomal status and cell viability. We chose to perform all experiments on non-capacitated sperm as this condition is the most consistent, less prone to donor-effects and is closer to the state in which the sperm will be when they would be encountering the male contraceptive drug. The goal was to be able to screen a 384-well plate in both assays consecutively within 90 min of sperm dispensing using the fully automatic robotic system. Data was analysed for inter- and intra-plate variation (both technical and biological replicates) and signal-to-background and variance measured to demonstrate the assay was reproducible and robust. The assay was shown to be scalable and all the workflows optimal. Once established, the system was validated by screening the ReFRAME library (Janes et al., 2018) and hits confirmed by dose-response analysis.
Sperm handling
Request a detailed protocolControl semen samples were obtained from volunteer donors with normal sperm concentration, motility and semen characteristics (WHO, 2010) and no known fertility problems. Samples were obtained by masturbation after 48–72 hr of sexual abstinence. After ~ 30 min of liquefaction at 37°C, cells were prepared using a discontinuous density gradient procedure (DGC). Semen was loaded on top of a 40–80% suspension of Percoll (Sigma Aldrich, UK) diluted with non-capacitation medium using Minimal Essential Medium Eagle, supplemented with HEPES, Sodium lactate and Sodium Pyruvate to achieve a similar buffer as described previously (Tardif et al., 2014). We routinely pooled samples after DGC from 3 to 5 donors for use in each screening batch run in order to reduce donor-to-donor variability. Samples were obtained and analysed in line with suggested guidance for human semen studies where appropriate (Björndahl et al., 2016).
Motility assay
Request a detailed protocolSperm cells were incubated for 3 hr at 37°C after DGC, transferred to the robotic platform (HighRes Biosolutions Inc) and maintained at 37°C with gentle stirring (100 rpm) in a water bath on top of a magnetic stirrer. Approximately 10,000 spermatozoa (10 μl) were dispensed per well using a MultiDrop Combi (ThermoFisher) into pre-warmed assay-ready plates. Spermatozoa were incubated with the compounds for 10 mins (thus favouring fast acting compounds) at 37°C in the CV7000 microscope prior to the commencement of imaging.
Time-lapse imaging
Request a detailed protocolA CV7000 Cell Voyager high-content imaging system (Yokogawa) was used as it allowed full environmental control, sufficient contrast using simple brightfield optics and a fast acquisition rate (up to 45 frames per second). Using a 20x lens with bright field illumination (0.11 ms exposure, 3% lamp power), time-lapse image series were acquired (24 frame in 0.5 s) at two positions per well (with a 400 μm gap between positions to eliminate double counting of sperm). Image acquisition across a 384-well plate with such settings takes ~ 17 min.
Sperm tracking
Request a detailed protocolA Python implementation of the particle tracking algorithm originally developed by others (Crocker and Grier, 1996) was employed (Trackpy v0.4.1 Allan, 2018). The algorithm determines the position of every sperm head in each frame and links the position over time creating tracks. We optimized the algorithm parameters for our imaging data to exclude sperm cross-tracks and avoid detecting non-sperm particles. The image data (~17,000 files amounting to 21 GB per plate) can be processed within 30 min using a standard desktop PC (Intel Core i5-6600 CPU, 3.3 GHz, 8 Gb RAM) and can also be parallelized on a compute cluster with minimal effort.
Acrosome assay
Request a detailed protocolA flow cytometry-based assay was developed employing an iQue Screener (Sartorius) with Alexa488-conjugated peanut agglutinin (PNA) (Mortimer et al., 1987) and general sperm viability (using propidium iodide, Pi). This assay was run directly after motility assessment. Controls (DMSO and the Ca2+ ionophore A23187) were added using an acoustic dispenser (Echo 555, Labcyte Inc) and incubated for 10 min at 37°C in the SteriStore (HighRes Biosolutions Inc). PNA488 (ThermoFisher Scientific, Cat. No. L21409, stored as 1 mg/mL stock) was then added to achieve a 1:1000 final dilution and propidium iodide (ThermoFisher Scientific, Live/Dead Sperm Viability kit, Cat. No. L7011, stored as 2.4 mM solution) was added to achieve a 1:2000 final dilution using a liquid handler (Tempest, Formulatrix). After addition of dyes, plates were incubated for 10 min at 37°C in the SteriStore before sampling using an iQue Screener (2 s sip time per well with pump setting of 45 rpm). This resulted in a sample processing time of about 25 min per 384-well plate. Cells were categorised as either: unstained cells; ‘dead’ cells (Pi+ PNA-); Acrosome-reacted but ‘dead’ cells (Pi+ PNA+); or Acrosome-reacted ‘live’ cells (Pi- PNA+). This method is capable of detecting a shift of populations (Pi+ and PNA+) upon induction of AR using A23187. The AR assay was performed as an agonist screen (scoring for induction of AR in live cells compared to DMSO controls) with the expectation that compounds could be found that induce AR beyond levels of induction with A23187.
Controls and QC criteria
Request a detailed protocolFor the motility assay DMSO was used as a vehicle control (negative control) and Pristimerin (Merck, Cat. No. 530070, stored as 10 mM stock in DMSO; final concentration of 20 µM) as a positive control. For the acrosome assay we added a calcium ionophore (A23187, Sigma-Aldrich, Cat. No. C7522, stored as 10 mM stock in DMSO, used at a final concentration of 10 µM) as a positive control and DMSO was added to wells as a negative control to be at the same final concentration as that present in the compound-containing wells (0.0625%). DMSO tolerance was tested by performing the standard motility assay exposing sperm to increasing concentrations (max 10%) of DMSO for 10 min. We calculate the standard statistical measure of assay robustness, the Z’ value (Zhang et al., 1999) for each plate using positive and negative controls for the motility assay and observed Z’ values ranging between 0.4–0.8. In addition, we performed a visual check of heatmaps for every plate to detect edge effects.
Data analysis and normalization
Motility
Request a detailed protocolcustom R scripts were written to calculate standard sperm kinetic parameters (Mortimer et al., 2015). Those parameters allow classification of sperm into standard WHO classes: progressively motile (PM) (where average path velocity: VAP > 25 um/sec AND straightness: STR > 80%); non-progressively motile (NPM) (where VAP > 5 um/sec OR straight line velocity: VSL > 11 um/sec), and immotile (IM). In addition to calculating kinetic parameters, we established a workflow to generate movies of time-lapse videos with overlapping sperm tracks using R and FFMPEG (FFmpeg Developers. Available from http://ffmpeg.org). We expressed results as % of control curvilinear velocity: VCL. This was defined as VCL_median (cpd)/VCL_median(DMSO)*100. Hit selection criteria was 15% reduction of VCL. VCL was chosen as the main kinetic measurement as it had an acceptable Z’ value and is independent of path averaging. Wells with autofocus errors or compounds which did not dissolve properly (resulting in sperm cells being stuck in one location but moving normally in the rest of the well) were excluded from analysis.
Acrosome
Request a detailed protocoliQue Screener Data was exported as FCS format and processed using the following Bioconductor packages for analysing flow cytometry data: flowCore, flowDensity, flowWorkspace, ggcyto (see https://bioconductor.org). These packages allow handling flow cytometry data as objects, which can then be compensated, gated and visualized efficiently. In addition, we use HDF5 format (www.hdfgroup.org) for storing processed and averaged data. Percent events for each population (Pi+ PNA+, Pi- PNA+, Pi+ PNA- and Pi- PNA-), normalized to total well events, were calculated. Hits were defined as compounds which induce AR beyond 15% (maximum level of induction achieved with positive control). Wells with irregularities (low in Pi+ or Pi+PNA+ population, or below 200 events) have been excluded from analysis.
Compound screening
Request a detailed protocolWe screened the ~12,000 compound ReFRAME (Repurposing, Focused Rescue, and Accelerated Medchem) library (Janes et al., 2018; and www.reframedb.org) supplied by CALIBR at the Scripps Institute. This unique library consists of bioactives and approved drugs that have been assembled from the literature, drug databases and by patent mining. Compounds dissolved in DMSO were spotted (12.5 nL, final assay concentration ~ 6 μM) into 384-well black-sided optical imaging plates (CellCarrier, PerkinElmer) at CALIBR and were shipped on dry ice to the screening centre in Dundee and stored at −20 °C until required. Immediately prior to screening plates were thawed, controls were added using the acoustic dispenser (Echo 555, Labcyte), and 10 μl warm media added followed by shaking on a plate shaker for 10 s. Plates were then incubated for 30 min at 37°C to solubilise the compounds and prewarm the plates prior to the addition of live sperm (see above). With the addition of the media containing sperm (10 μl) the final concentration of DMSO in each well was 0.0625%.
All experiments were performed with pools of donors (3–5 per batch of 4 screening plates) and results normalised to DMSO-treated wells (16 wells per plate). Sperm was first prepared by DGC and then pooled. Where a single donor sample was found to less than 1/3 of the yield required then the contribution from the other doors was increased to compensate.
The ReFrame library has been screened for cytotox (using CellTiterGlo) in HEK293 and HepG2 cells (see www.reframedb.org) with some of the hit compounds displaying activity in one or both assays (Figure 2—source data 1), information which helps inform compound progression decisions.
Dose response experiments
Request a detailed protocolAssay-ready 384-well CellCarrier plates with 8-point curves (10 μM highest concentration) were resupplied by CALIBR in duplicate and processed as described above. Curves were fitted using the R package dr4pl (https://cran.r-project.org/web/packages/dr4pl/index.html) using a 4-parameter logistic fit option.
Triaging of AR hits
Request a detailed protocolGiven the prevalence of a range of interfering compounds amongst the library hits, two approaches were used for triaging. These additional triaging steps, after the primary screen, allowed for the elimination of intrinsically fluorescent compounds and compounds fluorescing in the presence of biological material. Screening plates were excited at 488 nm and read at 520 nm and 670 nm in a M1000Pro multimode plate reader (Tecan) after flow cytometry. In addition, when performing dose-response curve follow up on resupply material a replicate set of compound-dosed cells were processed with no-dyes added. This identified the small number of compounds whose fluorescence is indistinguishable from a true positive. In addition, visual confirmation of the appropriate pattern of acrosome staining was performed by microscopy.
Disulfiram washout assay
Request a detailed protocolIn order to assess the ability of sperm cells to recover motility after Disulfiram removal, wash-out experiments were conducted at two timepoints using non-capacitating conditions. Spermatozoa were pooled after DGC preparation in the same manner as for screening. Pooled samples were diluted to a concentration of approximately 106/mL and incubated at 37°C under non-capacitating conditions (see above). After this initial incubation period, Disulfiram (10 μM final concentration) or vehicle control (DMSO to 0.1% final concentration) were added and incubated at 37°C for 20 min. Following exposure to Disulfiram or DMSO control, sperm cells were washed by centrifugation at 300 g for 5 min in 5 mL of the non-capacitation media. After washing, the pellet was resuspended in 500 μl of the appropriate media and incubated at 37°C for 1 hr. Samples were taken for analysis by computer-assisted-sperm analysis (CASA) [CEROS machine (version 12), Hamilton Thorne Research, Beverly, MA, USA] before treatment, after 20 mins treatment and 1 hr post washout. Wash-out experiments were performed on three separated days, utilising different donors to make up the pooled sample. Each condition was repeated in duplicate on each of these days and each CASA reading was performed in duplicate.
Data availability
Full data is available. Large files have been deposited to Dryad (http://doi.org/10.5061/dryad.jdfn2z36z).
-
Dryad Digital RepositoryData from: A phenotypic screening platform utilising human spermatozoa identifies compounds with contraceptive activity.https://doi.org/10.5061/dryad.jdfn2z36z
References
-
Data Analysis, Machine Learning and Applications319–326, KNIME: The Konstanz Information Miner, Data Analysis, Machine Learning and Applications, Springer, 10.1007/978-3-540-78246-9_38.
-
Quantifying the chemical beauty of drugsNature Chemistry 4:90–98.https://doi.org/10.1038/nchem.1243
-
Methods of digital video microscopy for colloidal studiesJournal of Colloid and Interface Science 179:298–310.https://doi.org/10.1006/jcis.1996.0217
-
ReportMen as Contraceptive Users: Programs, Outcomes and RecommendationsWashington, DC: Population Council, The Evidence Project.
-
Role of disulfide linkage in action of bis(dialkylaminethiocarbonyl)disulfides as potent double-Edged microbicidal spermicide: design, synthesis and biologyEuropean Journal of Medicinal Chemistry 115:275–290.https://doi.org/10.1016/j.ejmech.2016.03.012
-
CatSper: a unique calcium channel of the sperm flagellumCurrent Opinion in Physiology 2:109–113.https://doi.org/10.1016/j.cophys.2018.02.004
-
Opportunities and challenges in phenotypic drug discovery: an industry perspectiveNature Reviews Drug Discovery 16:531–543.https://doi.org/10.1038/nrd.2017.111
-
The future of computer-aided sperm analysisAsian Journal of Andrology 17:545–553.https://doi.org/10.4103/1008-682X.154312
-
Male hormonal contraceptive - are we there yet?Nature Reviews Endocrinology 14:685–686.https://doi.org/10.1038/s41574-018-0111-4
-
RDKit: Open-Source Cheminformatics and Machine LearningRDKit: Open-Source Cheminformatics and Machine Learning, http://www.rdkit.org.
-
Reproductive endocrinologists are the gatekeepers for male infertility care in north America: results of a north american survey on the referral patterns and characteristics of men presenting to male infertility specialists for infertility investigationsFertility and Sterility 112:657–662.https://doi.org/10.1016/j.fertnstert.2019.06.011
-
Investing in family planning: key to achieving the sustainable development goalsGlobal Health: Science and Practice 4:191–210.https://doi.org/10.9745/GHSP-D-15-00374
-
How were new medicines discovered?Nature Reviews Drug Discovery 10:507–519.https://doi.org/10.1038/nrd3480
-
BookWHO Laboratory Manual for the Examination and Processing of Human Semen (Fifth)Cambridge: Cambridge University Press.
-
Prediction of physicochemical parameters by atomic contributionsJournal of Chemical Information and Computer Sciences 39:868–873.https://doi.org/10.1021/ci990307l
-
A simple statistical parameter for use in evaluation and validation of high throughput screening assaysJournal of Biomolecular Screening 4:67–73.https://doi.org/10.1177/108705719900400206
Article and author information
Author details
Christopher LR Barratt

Funding
Bill and Melinda Gates Foundation (OPP1160989)
- Franz S Gruber
- Christopher LR Barratt
- Paul D Andrews
Bill and Melinda Gates Foundation (OPP1203050)
- Franz S Gruber
- Zoe C Johnston
- Christopher LR Barratt
- Paul D Andrews
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication. The funders facilitated using the REFRAME library
Acknowledgements
We are very grateful to all members of the research team for their invaluable assistance. We also want to thank all the sperm donors who took part in this study and Dr Stephen Gellatly and Ms Morven Dean for recruitment. We acknowledge the assistance of Irene Sucquart in the early phase of this study. Additionally, we want to thank Dr David Mortimer and Dr Sharon Mortimer for their helpful insights into comparisons with the CASA system and advice in AR assay development. Thanks go to Dr Steve Publicover and Professor C De Jonge for critical reading of the manuscript. Thanks are also due to NPSC lab members for help, particularly John Raynor for engineering support. We thank Mitch Hull, Emily Chen and Kelli Kunen at CALIBR for their help in library plating, logistics and supply of ReFRAME data. Thanks also go to Professor Kevin Reid, Professor Ian Gilbert and Dr Caroline Wilson of Drug Discovery Unit in Dundee for helpful discussions. Special thanks are due to Professor Andrew Hopkins (AH) for his support.
Ethics
Human subjects: Written consent was obtained from each donor in accordance with the Human Fertilization and Embryology Authority (HFEA) Code of Practice (version 8) under local ethical approval (13/ES/0091) from the Tayside Committee of Medical Research Ethics B.
Copyright
© 2020, Gruber et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 3,819
- views
-
- 528
- downloads
-
- 21
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 21
- citations for umbrella DOI https://doi.org/10.7554/eLife.51739
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A phenotypic screening platform utilising human spermatozoa identifies compounds with contraceptive activity
eLife 9:e51739.
https://doi.org/10.7554/eLife.51739




{kind=link}
{kind=link}
{kind=link}