Cystic proliferation of germline stem cells is necessary to reproductive success and normal mating behavior in medaka
- Instituto Tecnológico de Chascomús, INTECH (CONICET-UNSAM), Argentina
- Reproductive and Molecular Biology Group, Department of Structural and Functional Biology, Institute of Biosciences, São Paulo State University (UNESP), Brazil
- Salmonid Experimental Station at Campos do Jordão, UPD-CJ, Sao Paulo Fisheries Institute (APTA/SAA), Brazil
Abstract
The production of an adequate number of gametes is necessary for normal reproduction, for which the regulation of proliferation from early gonadal development to adulthood is key in both sexes. Cystic proliferation of germline stem cells is an especially important step prior to the beginning of meiosis; however, the molecular regulators of this proliferation remain elusive in vertebrates. Here, we report that ndrg1b is an important regulator of cystic proliferation in medaka. We generated mutants of ndrg1b that led to a disruption of cystic proliferation of germ cells. This loss of cystic proliferation was observed from embryogenic to adult stages, impacting the success of gamete production and reproductive parameters such as spawning and fertilization. Interestingly, the depletion of cystic proliferation also impacted male sexual behavior, with a decrease of mating vigor. These data illustrate why it is also necessary to consider gamete production capacity in order to analyze reproductive behavior.
Introduction
Cell division and differentiation are well-controlled processes for the development of a functional organ in multicellular organisms. In gonads, these processes are also essential to guarantee the correct production of gametes necessary for successful reproduction (Lombardi, 1998). Vertebrate gonads acquire sex-specific features during gonadogenesis, an exceptional, complex, multistep process that includes migration of primordial germ cells (PGCs) to reach the gonadal primordium, proliferation, differentiation, and meiosis (DeFalco and Capel, 2009; Richardson and Lehmann, 2010). Defects in any of these steps may lead to sex abnormalities including infertility and sex reversal.
Proliferation of embryonic germline stem cells (EGSCs, the name applied to PGCs after they reach the gonadal primordium) gives rise to a cohort of germ cells that will ultimately form the primary population of germ cells in the gonads of both sexes (Tanaka, 2019). This type of slow, intermittent division, also called type I proliferation, is similar to the self-renewal of germline stem cells in the adult ovary or testis (Chen and Liu, 2015; Nakamura et al., 2010), and is responsible for ensuring the production of the correct number of germline stem cells that will initiate gametogenesis (Tanaka, 2019). In addition, the hyperproliferation of mitotically active germ cells in hotei medaka, a mutant fish for anti-Müllerian hormone receptor II (amhrII), induces 50% male-to-female sex reversal and loss of reproductive success in adult fish (Morinaga et al., 2007; Nakamura et al., 2012). On the contrary, lack of this proliferation of EGSCs during the embryo stage induces the opposite sex reversal in zebrafish and medaka (Kurokawa et al., 2007; Rodríguez-Marí et al., 2010), showing that this initial proliferation can be also involved in gonadal sex fate.
A second EGSC proliferation occurs prior to meiosis, following a special program of synchronous, successive and incomplete mitosis, generating interconnected daughter cells known as cysts or nests, which are surrounded by somatic cells (Saito et al., 2007). Following several rounds of mitosis, the cells enter meiosis to become oocytes in the ovary or spermatocytes in the testis (Lei and Spradling, 2013b; Matova and Cooley, 2001; Tanaka, 2016). Cystic division is a highly conserved mechanism that precedes gametogenesis in both invertebrates and vertebrates (Amini et al., 2014; Hansen and Schedl, 2006; Hinnant et al., 2017; Lei and Spradling, 2013a; Quagio-Grassiotto et al., 2011). It has been well established in vertebrates that female germ cells enter meiosis earlier, while male germ cells are arrested and undergo meiosis at the onset of the prepubertal period (Elkouby and Mullins, 2017; Koubova et al., 2006; Lei and Spradling, 2013a; Pepling, 2006). In medaka, this type of cystic cell division is known as type II proliferation, by which, in females, the number of EGSCs increases dramatically during embryonic stages; whereas in males, this exponential proliferation to form cysts begins later, in the juvenile stage (Satoh and Egami, 1972; Tanaka, 2019). Despite the importance of cystic cell proliferation during early life to ensure reproductive success, this process is not fully understood at the molecular level (Elkouby and Mullins, 2017).
In the search for genes that regulate EGSCs proliferation, ndrg1 (n-myc downstream regulated gene 1) is one of the genes that was found to be upregulated in germ cells with mutated foxl3, a switch gene involved in the germline sexual fate decision in medaka (Kikuchi et al., 2019). Interestingly, Ndrg1 is involved in the regulation of cell proliferation in different cancers (Chang et al., 2014; McCaig et al., 2011; Xi et al., 2017; Zhang et al., 2019). Moreover, this gene has homologs from mammals (Ndrg1) to fish (ndrg1), Caenorhabditis elegans (ZK1073.1) to fruit fly (Dmel\MESK2), suggesting that NDRG1 is highly conserved among species (Kovacevic et al., 2013; Sun et al., 2013). Ndrg1 has been demonstrated to inhibit the TGF-β signaling cascade, a very well-known pathway involved in cell proliferation and survival (Zhang et al., 2017), regulating the induction of epithelial-to-mesenchymal transition in mammalian cells (Tojo et al., 2005; Chen et al., 2012; Jin et al., 2014), establishing ndrg1 as a potent inhibitor of proliferation. However, its role in non-cancerous cells is not well established. In this study, we provide clear evidence of the important role of ndrg1 in the regulation of EGSC cystic proliferation, with subsequent impact on reproductive success in medaka.
Results
Expression of ndrg1 during gonadal development
To understand the role of ndrg1 in cystic proliferation, the expression pattern of ndrg1 during gonadal development was first determined. ndrg1 belongs to the n-myc downstream regulated gene 1 family, and in the medaka genome has two paralogs (Figure 1A), as a result of teleost-specific genome duplication (Kasahara et al., 2007): ndrg1a (ENSORLG00000003558) on chromosome 11 and ndrg1b (ENSORLG00000004785) on chromosome 16, with 60% identity. The ndrg1a transcripts were not detectable in the medaka gonads during embryo developmental stages by whole-mount RNA in situ hybridization (Figure 1B). In contrast, the medaka ndrg1b was specifically observed in gonads during embryo developmental stages (Figure 1C). The transcript level of ndrg1b was quantified by qPCR at stage 35, when female (XX) and male (XY) gonads appear morphologically indistinguishable and germ cells only show type I proliferation. No difference for ndrg1b transcript levels was detected between female and male individuals (Figure 1D). Interestingly, differences between sexes were observed at stage 39, at 10 and 20 days post hatching (dph) (Figure 1E–G). At stage 39 and 10 dph, where female gonads have higher number of cysts compared to male gonads due to type II cystic division, the level of ndgr1b transcripts is higher in male than in female gonads (Figure 1E, F). The opposite sex transcriptional profile of ndrg1b was observed at 20 dph (Figure 1G), when type II proliferation begins in male individuals. These results showed that the transcription of ndrg1b is inversely related to the cystic proliferation of EGSCs in both sexes.
Figure 1 with 1 supplement see all
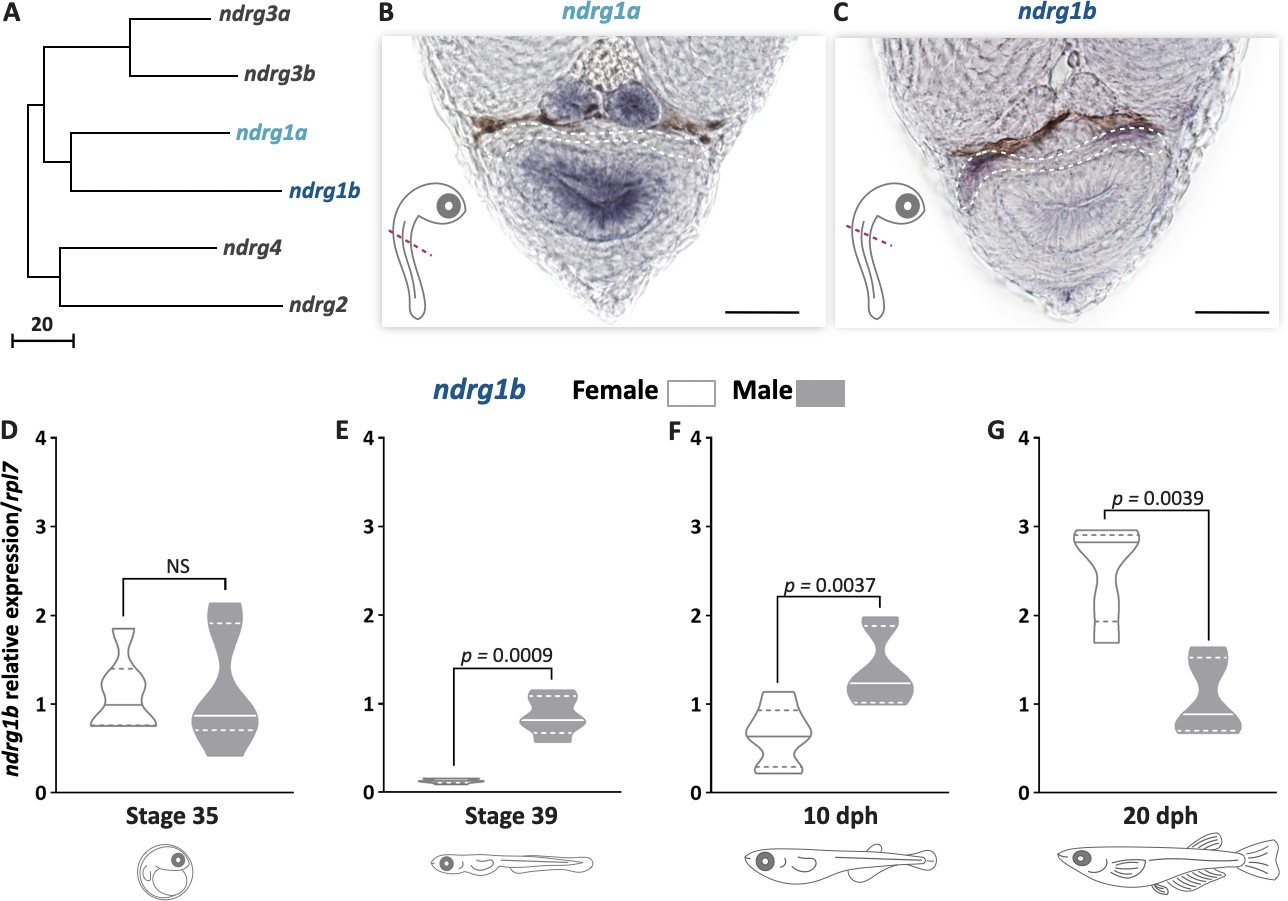
ndrg1b is down-regulated during cystic proliferation in gonadal development.
Phylogenetic tree showing the relationships among the ndrg family members in medaka (A), obtained using the neighbor-joining method and a bootstrap test (MEGA 7.0 software). The scale beneath the tree reflects sequence distances. ENSEMBL accession sequences are provided in Supplementary file 1A. Transversal sections of gonadal region (red line) in whole embryos at stage 35 to detect ndrg1a (B) and ndrg1b (C) transcripts using in situ hybridization. Gonadal region is surrounded by white dotted line; scale bar represents 20 µm. Transcript abundance levels of ndrg1b in different stages of gonadal development: 35 (D), 39 (E), 10 days post hatching (dph) (F), and 20 dph (G). Quantification was performed using the 2-ΔΔCt method and ndrg1b values were normalized to rpl7. Genotypic sex was determined by the presence/absence of the dmy gene; female (XX) and male (XY) are represented by empty bars or full bars, respectively. n = 5–6 pools per sex in stages 35 and 39, and n = 6–7 individuals per sex in 10 and 20 dph. p-Values are indicated when transcript abundance between sexes at the same developmental stage differ significantly (p<0.05). NS: not statistically significant. Relative gene expression levels were compared as described by Pfaffl, 2001.
TGF-β pathway during proliferation and its relation with ndrg1b
Due to the key role of the TGF-β superfamily in the regulation of type I proliferation in medaka (Imai et al., 2015; Nakamura et al., 2012) and the effect of ndrg1b on that proliferation, we decided to analyze whether TGF-β signaling is involved in the regulation of cystic proliferation (Figure 1—figure supplement 1B). Embryos of both sexes were exposed to A83-01 (TGF-β inhibitor), from stage 35 to 39 (Figure 1—figure supplement 1A). When total EGSCs were quantified, in exposed females no difference was observed, but an increase in EGSCs in exposed males was observed compared to control males (Figure 1—figure supplement 1C). Additionally, no difference in ndrg1b mRNA levels was detected between control and treatment for female individuals (Figure 1—figure supplement 1D). In contrast, male individuals showed increased levels of ndrg1b following TGF-β inhibitor treatment when compared to control (Figure 1—figure supplement 1D), establishing that there is no negative feedback from ndrg1b via TGF-β. Overall, these results suggest that the TGF-β pathway is not involved in type II proliferation in medaka.
Mutation of ndrg1b affects early gonadal development in both sexes
To explore the role of ndrg1b during gonadal development and cystic proliferation, firstly ndrg1b biallelic mutants (indels in F0 individuals genome created by injecting Cas9 and sgRNA) were generated using CRISPR/Cas9 technology. The mutagenesis efficiency for each sgRNA was analyzed by a heteroduplex mobility assay (HMA; Figure 2—figure supplement 1B, Figure 2—figure supplement 2B; Ota et al., 2013), which reached 96.6% for sg1_ndrg1b and 80% for sg2_ndrg1b (Figure 2—figure supplement 1B, Figure 2—figure supplement 2B). These results led us to select sg1_ndrg1b for subsequent analysis. In addition, biallelic mutants were mated with wild-type (wt) fish to generate F1 and then several fish were sequenced to confirm the presence of the indels (Figure 2—figure supplement 1C). The data indicate that most cells contained biallelic indels and, consequently, loss of function in ndrg1b mutants. None of the embryos analyzed had indels at the analyzed off-target sites for the injected sgRNA of biallelic mutant (Figure 2—figure supplement 1D, E). In order to address the possible involvement of ndrg1b in sex development, biallelic mutants (F0) were analyzed.
Fish having the deletion Δ31 (deletion of 31 bp) previously identified (Figure 2—figure supplement 1C) were selected as founder F1 to produce the F2 generation with expected ndrg1b knockout (KO) (Figure 2—figure supplement 3A). However, the screening of F2 individuals by means of HMA using genomic DNA from a fin clip (Figure 2—figure supplement 3C, D) did not reveal homozygous mutants of ndrg1b (Figure 2—figure supplement 3B).
Mutation of ndrg1b affects early gonadal development in both sexes
To analyze the phenotypic alterations of gonad development in ndrg1b mutants, the number of EGSCs was counted using the germ cell marker Olvas at stage 39 (Figure 2A–C). The total number of EGSCs in male sgN1b was unchanged with respect to the male wt individuals. In females, however, the number was significantly reduced in female sgN1b compared to the female wt individuals (Figure 2D), decreasing to a level similar to male wt. Moreover, when the number of type I and II germ cells in mutants was quantified, only type II cystic proliferation was altered in female mutants compared with wt female individuals (Figure 2F). Furthermore, the number of EGSCs inside each cyst in sgN1b females was lower than in wt (Figure 2E).
Figure 2 with 4 supplements see all

The mutation of ndrg1b affects gonadal development at stage 39.
Schematic representation of the experimental procedure adopted to analyze the effect of ndrg1b loss on germ cell number and proliferation (A: anterior, P: posterior, D: dorsal; and V: ventral) (A). Fluorescent images of gonads in female or male embryos; wild-type (wt) = injected with cas9 (B) and sgN1b = injected cas9+sg1_ndrg1b (C). Embryonic germline stem cells (EGSCs) were stained with an anti-OLVAS antibody (green), nuclei were stained with DAPI (blue), and proliferating cells with PCNA (: Proliferating cell nuclear antigenDMSO: Dimethyl sulfoxide) antibody (red), scale bar represents 20 µm. Total numbers of EGSCs (D), number of EGSCs per cyst with type II division (E), total numbers of germ cells with type I or type II division (F), and number of PCNA-positive type I and cystic type II germ cells relative to the total number of type I or type II germ cells, respectively (G), in wt female or male embryos (represented by gray empty bars or full bars, respectively) and sgN1b female or male embryos (represented by cyan empty bars or full bars, respectively). Vertical bars indicate mean, with its respective standard error of the mean. n = 8 per each wt group and n = 12 per each sgN1b group. p-Values are indicated when the number of germ cells between groups differs significantly (p<0.05). NS: not statistically significant. Unpaired Student’s t-test per E and Tukey’s multiple comparisons test per D, F, and G.
We then wondered whether the loss of EGSCs was due to reduced cystic proliferation or to apoptosis in the ndrg1b mutants. PCNA immunostaining (Figure 2B, C) revealed that the number of proliferating type I and type II germ cells/total number of germ cell at stage 39 did not differ among female sgN1b and male gonads, but the number of proliferating type II germ cells was reduced in female sgN1b when compared to female wt (Figure 2G). In a TUNEL assay, positive cells were not observed in any sgN1b or wt individuals (Figure 2—figure supplement 4A–C). Hence, female ndrg1b mutants have fewer proliferating germ cells at stage 39. These results indicate that type II cystic proliferation is affected in ndrg1b mutants. In addition, we performed an extra check of the efficacy of the loss of function of ndrg1b with the sgN1b guide by analyzing the phenotype of loss of cystic proliferation in individuals injected with a second RNA guide, sg2_ndrg1b. We observed the same results as the sgN1b biallelic mutants (Figure 2—figure supplement 2C–E). These results confirm the ndrg1b loss of function observed with the first sgRNA.
The effect of ndrg1b mutation was further analyzed during juvenile development in both sexes at 20 dph, which is the time when type II cystic division begins in males. Histological examination showed ovaries with apparently normal development (Figure 3B); however, there is a decrease in the number of oogonia and pre-vitellogenic oocytes compared to wt individuals (Figure 3D, E). In males, the mutant showed a significant reduction of cysts and germ cells within the cyst in relation to wt males (Figure 3F, G). This indicates that germ cell number is affected in ndrg1b mutants for both sexes at times that are key to the commitment of type II cyst division.
Figure 3
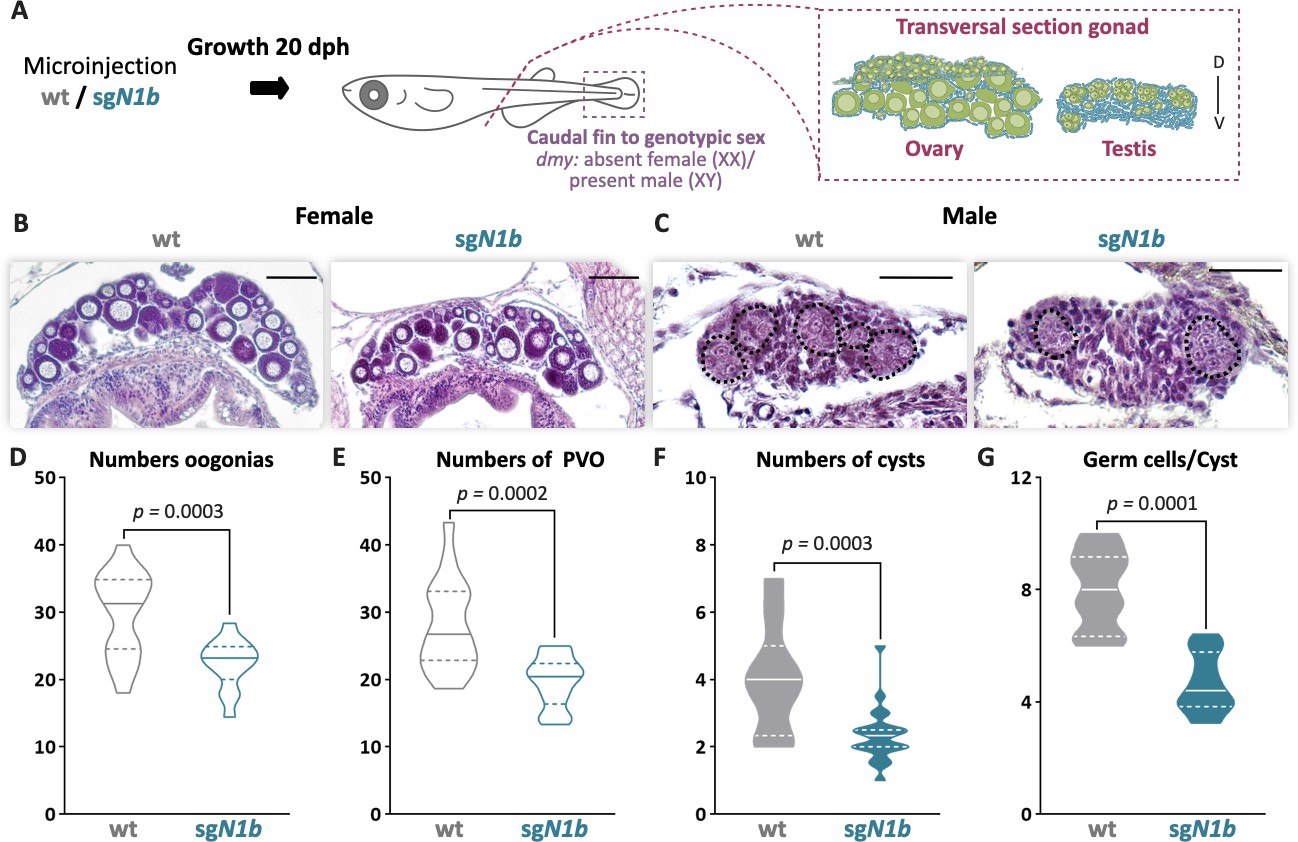
The mutation of ndrg1b affects gonadal development at 20 days post hatching (dph).
Schematic representation of the experimental procedure adopted to analyze the effect of ndrg1b mutation on gonadal histology at 20 dph (D: dorsal, V: ventral) (A). Histological transverse sections of gonads. Wild-type (wt) = injected with cas9 or sgN1b=injected with cas9+sg1_ndrg1b from individuals for XX (B) and XY (C). In the testis, each cyst of germ cells (spermatogonia-like cells) is encircled by a black dotted line. Scale bars in B and C represent 50 µm. Total number of oogonia (D), number of pre-vitellogenic oocytes (PVO) (E), number of spermatogonial cysts (F), and number of spermatogonia per cyst (G) in wt female or male embryos (represented by gray empty bars or full bars, respectively) and sgN1b female or male embryos (represented by cyan empty bars or full bars, respectively). n = 16 per female groups and n = 20 per male groups. p-Values are indicated when the number of germ cells between groups differs significantly (p<0.05). NS: not statistically significant. Unpaired Student’s t-test.
Mutation of ndrg1b alters gametogenesis and reproductive success of both sexes in adulthood, but not androgen production in males
To evaluate whether impaired type II cystic proliferation can affect gametogenesis and reproductive success, paired adult male and female wt and ndrg1b mutants were analyzed for 10 days, and then their body and gonadal morphology was analyzed according to different reproductive parameters (Figure 4A). Regarding the daily frequency of spawning, the ndrg1b mutant females spawned fewer eggs than wt females (Figure 4B), although fertilization rate was normal (Figure 4C). Surprisingly, the wt females paired with male sgN1b also spawned fewer oocytes (Figure 4B), and in sgN1b males paired with wt females, the fertilization rate declined (Figure 4C). Moreover, hatching rate was lower for the progeny of male sgN1b and wt female than for the progeny of female sgN1b and wt male (Figure 4D). Similar results were observed in F1 (heterozygote Δ31), in which the number of spawning eggs was reduced in female and male ndrg1b+/- (Figure 4—figure supplement 1A), the fertilization rate was reduced in males (Figure 4—figure supplement 1B), and, interestingly, the hatching rate was sex dimorphic, reduced only in embryo from males compared to the respective control male (Figure 4—figure supplement 1C), indicating a need for future studies in this direction.
Figure 4 with 1 supplement see all
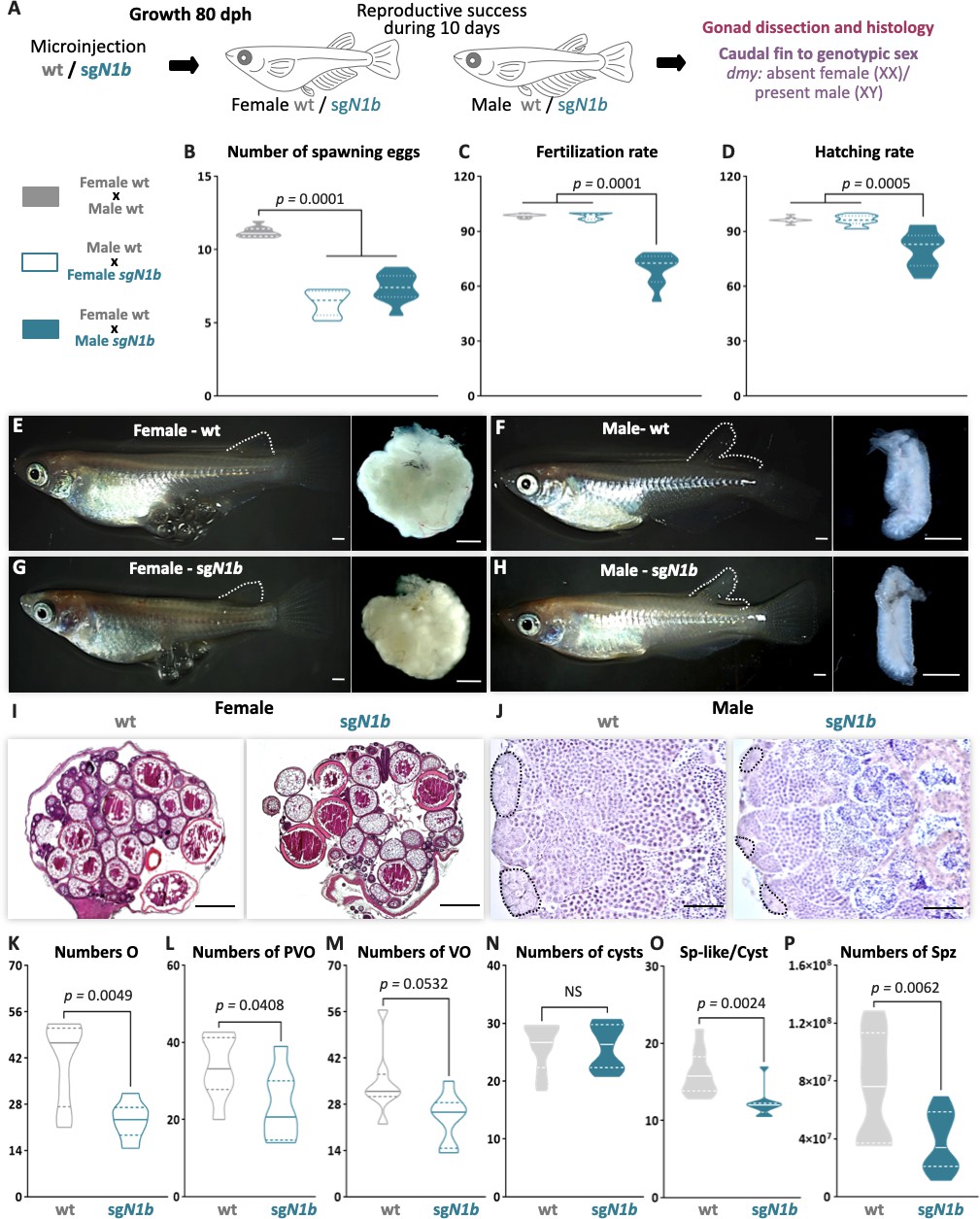
Mutation of ndrg1b alters the gametogenesis and reproduction success of both sexes in adults.
Schematic representation of the experimental procedure adopted to analyze the effect of ndrg1b loss of function in the reproductive success at 80 days post hatching (dph) (A). Reproductive success was evaluated by crossing the wild-type (wt) males with sgN1b females (cyan full), wt females with sgN1b males (cyan empty), and as a control, wt females with wt males (gray). For all crossings, we quantified the total number of eggs spawned (B), percentage of fertilization (C), and percentage of hatching (D) of the total eggs spawned. The biallelic mutants of ndrg1b did not exhibit morphological sexual differences: adult females with their ovaries, wt (E) and sgN1b (G), and adult males with their testis, wt (F) and sgN1b (H). The secondary sexual characteristics are indicated by the separation of hindmost rays from other rays in the dorsal fin. Scale bars are 1 mm. Histological transverse sections of wt and sgN1b gonads from XX (I) or male (J) individuals. In the testis, each cyst of germ cells (spermatogonia-like cells) is encircled by a black dotted line. Scale bars represent 500 µm (I) and 50 µm (J). Number of oogonia (O) (K), number of pre-vitellogenic oocytes (PVO) (L), number of vitellogenic oocytes (VO) (M), number of spermatogonial cysts (N), number of spermatogonia per cyst (O), and number of spermatozoa (P) from wt female or male individuals (represented by gray empty bars or full bars, respectively) and sgN1b female or male individuals (represented by cyan empty bars or full bars, respectively). n = 10 per group. p-Values are indicated when groups differ significantly (p<0.05). NS: not statistically significant. Tukey´s multiple comparisons test (B–D). Unpaired Student’s t-test (K–P).
Medaka adults display easily distinguishable secondary sex characteristics, for example, the hindmost rays are separated from other rays in the dorsal fin in the male, but linked together in female. All ndrg1b mutants invariably displayed typical female and male secondary sex characteristics (Figure 4E–H). A balanced sex ratio was observed in both ndrg1b mutants and wt individuals (Supplementary file 1B). Histological examination of females showed ovaries with apparently normal development (Figure 4I), but there were significantly fewer oogonia, pre-vitellogenic, and vitellogenic oocytes at the different stages in ndrg1b mutants than in wt individuals (Figure 4K–M). With respect to the histological examination of testis, all stages of spermatogenesis were observed in sgN1b mutants (Figure 4J). Spermatogonia are surrounded by Sertoli cells at the distal end of the lobule, which are responsible for constantly supplying new germ cells by proliferation (Iwai et al., 2006). When these cells were counted manually, the number of cysts containing spermatogonia remained unchanged (Figure 4N); however, the number of spermatogonia within each cyst was lower in sgN1b than in wt individuals (Figure 4O), and the total number of spermatozoa was lower in sgN1b testes than in wt testes (Figure 4P). Collectively, these results indicate that alterations of cystic proliferation, with the concomitant lower sperm count and fewer oocytes, have direct influence on reproduction. Interestingly, these alterations do not change the general gonadal morphology or secondary sex characteristics in either sex or the sex ratio.
Mutation of ndrg1b disturbs the mating behavior of males
To explore a possible cause of the lower spawning of control females paired with male ndrg1b mutants, our next step explored the sexual behavior of these couples. Fortunately, the sexual behavior of medaka consists of a sequence of actions that are easily quantified (Hiraki-Kajiyama et al., 2019; Ono and Uematsu, 1957; Walter and Hamilton, 1970). Briefly, the sequences begin with the male approaching and following the female closely. The male then performs a courtship display, in which the male swims quickly in a circular pattern in front of the female. If the female is receptive, the male grasps her with his dorsal and anal fins (termed ‘wrapping’), they quiver together (termed ‘quivering’), and the male makes several gentle taps, like ‘convulsions’ (termed ‘convulsions’), and then oocytes and sperm are released. In general, all couples displayed the usual actions of sexual behavior (Figure 5B–I), with differences observed only in the actions related to mating behavior, particularly in the wrapping actions related to the stimulation of the female spawning, the duration of quivering, and number of convulsions, which were significantly fewer in sgN1b males paired with wt females than in control couples (Figure 5H, I). Collectively, these results indicate that alterations of cystic proliferation, with the concomitant lower sperm count, have direct influence on reproduction and mating behavior.
Figure 5 with 2 supplements see all
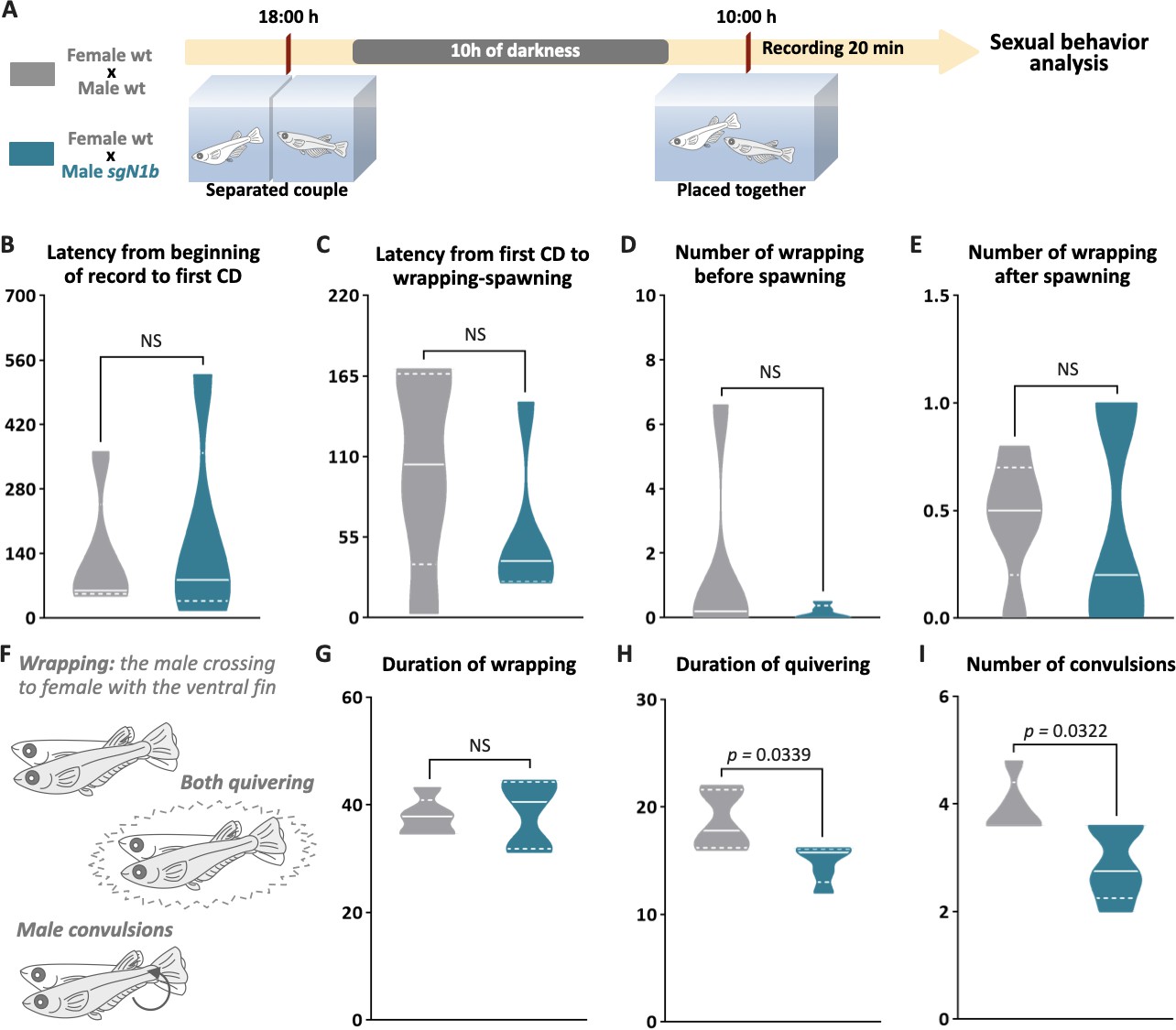
Reduction of cystic proliferation in ndrg1b mutants decreases male sexual behavior, with a decrease of mating vigor.
Schematic representation of the experimental procedure adopted to analyze reproductive behavior for 5 days; five couples per group were analyzed. (A) Couples of wild-type (wt) individuals (gray) or sgN1b males with wt females (cyan) were separated in the evening (6–7 PM) the day before the assay using a transparent plastic tank with small holes for water exchange. The following morning, each mating pair was placed together in a single tank and sexual behavior was recorded for 20 min. Each video was analyzed to determinate the latency from beginning of recording to the first courtship display (B), latency from first courtship display to the wrapping that resulted in spawning (C), number of wrappings of 2 s of duration before (D) and after (E) spawning, and specific wrapping actions (F), such as duration of wrapping (G), duration of quivering (H), and number of convulsions (I). Representative videos of wrapping actions in couples of wt female and male (https://www.youtube.com/watch?v=KrI8t90_tMA&feature=youtu.be) and male sgN1b with female wt (https://www.youtube.com/watch?v=MCjhYG7lfwM&feature=youtu.be). n = 5 per group. p-Values are indicated when the number of germ cells between groups differs significantly (p<0.05). NS: not statistically significant. Unpaired Student’s t-test.
Although we observed that the mutated individuals did not present changes in secondary sexual characteristics, such as the separation of hindmost rays from other rays in the dorsal fin (Figure 4E–H), which is strongly related to the presence of androgens in males (Ngamniyom et al., 2009), the corroboration of androgen levels is necessary because they are related to reproductive behavioral changes in adults of medaka (Hiraki-Kajiyama et al., 2019). The 11-ketotestosterone (11-KT) quantification did not show differences between wt and sgN1b males in a reproductive trial (Figure 5—figure supplement 1A, B), supporting that reduction in cystic proliferation and a reduced number of germ cells in males would not affect sex steroid-producing somatic cells.
Considering the unchanged levels of androgens in the ndrg1b mutants but significant changes in reproductive behavior, we proceeded to analyze whether this gene is expressed in other extra-gonadal tissues, specifically in the brain. We observed that the expression of ndrg1b is restricted to the gonad (Figure 5—figure supplement 2A, B), not being observed in the brain (Figure 5—figure supplement 2A, E). Within the testis, ndrg1b was observed in presumptive A-type undifferentiated spermatogonia (SGa; Figure 5—figure supplement 2B) since the cells that presented ndrg1b marking are situated in the most peripheral region of the tubule, where oct4-positive cells (Figure 5—figure supplement 2C) or SGa are observed (Figure 5—figure supplement 2D).
Discussion
Ndrg1 homologs have been found in different species of invertebrates and vertebrates (Fang et al., 2014; Kovacevic et al., 2013; Sun et al., 2013). In mammals, it has been well established that Ndrg1 exerts an inhibitory effect on cancer cell proliferation (Chang et al., 2014; McCaig et al., 2011; Xi et al., 2017; Zhang et al., 2019). Additionally, several studies have shown that Ndrg1 is expressed in the gonads of humans, mice, sheep, and medaka (Burl et al., 2018; Kikuchi et al., 2019; Lachat et al., 2002; Qu et al., 2019; Stévant and Nef, 2019); however, its role in gonadal function and reproduction is still unknown. In medaka, there are two paralogs of ndrg1: ndrg1a and ndrg1b, although only ndrg1b is expressed in the gonad. Interestingly, ndrg1b expression is opposite to EGSC cystic proliferation in both sexes. Present from invertebrates to vertebrates, cystic division is a highly conserved mechanism that precedes gametogenesis and is crucial to reproductive success. However, its molecular regulation remains unknown (Dinardo et al., 2011; Hinnant et al., 2017; Narbonne-Reveau et al., 2006).
To study the participation of ndrg1b in cystic proliferation, our first idea was to generate the loss of function of this gene through the generation of a homozygous mutant. However, obtaining a mutant line for ndrg1b was not possible because in homozygosis it would appear to be lethal. From the analysis of F2 individuals, and waiting for the presence of 25% homozygous KOs to ndrg1b, surprisingly, this genotype was not obtained because individuals died during embryonic early development for unknown reason. Moreover, it is important to highlight that obtaining these F1 and F2 required a significant investment of time given the very low spawning rate of the heterozygote females, the low fertility of heterozygote males, and the low hatching rate. The impossibility of obtaining a stable line for the loss of function of ndrg1b led us to search for another robust and reproducible alternative.
In fish, and especially in medaka (Ansai and Kinoshita, 2014), it has been well established that the complex Cas9/sgRNA generated a high-efficiency biallelic mutations of injected animals (cas9+sgRNA). The high efficiency of biallelic mutant and the use of this approach to research in medaka were already corroborated recently in our laboratory (Castañeda Cortés et al., 2019). This high efficiency is observed in the present work too, in which two different sgRNAs used to generate the biallelic mutant of ndrg1b presented 96.6% for sg1_ndrg1b and 80% for sg2_ndrg1b. Despite this high efficiency, this could not translate into a robust and reproducible phenotype, so were performed different controls. In this regard, we analyzed the gonadal phenotype of a second sgRNA (sg2-ndrg1b) injection, obtaining the same reduction of cystic proliferation of germ cells and adding to the verification of the absence of off-target, it supports the high directionality of the CRISPR/Cas9 system to generate the loss of function through the generation of biallelic mutants for ndrg1b. Another important concern to analyze is the reproducibility of the chosen system. On this regard, the phenotype throughout the different experiments, both in embryonic, juveniles, and in adult stages, carried out in many different individuals, supports the high repeatability and robustness of the phenotype of the biallelic mutant of ndrg1b. Furthermore, in a previous study by Castañeda Cortés et al., 2019 in medaka using biallelic mutants of two different sgRNAs targeting other loci, the corticotropin-releasing hormone receptors (crhr1 and crhr2), no relevant gonadal phenotype was observed in biallelic mutant fish reared at 24°C, showing the lack of a relevant phenotype in the germline after the use of CRISPR/Cas9. In addition, and something that is a strong point of the present work, this phenotype of the biallelic mutant was also analyzed and observed in both sexes, which even present different developing times in cystic proliferation.
In the present study, we generated ndrg1b F0 mutants by using CRISPR/Cas9 technology and showed that the lack of ndrg1b leads to a reduction in the total number of germ cells by inhibiting cystic proliferation in both sexes during early gonadal development and adulthood. This is evidence that ndrg1b is likely involved in the molecular regulation of cystic division in medaka. Gametogenesis commences within the spermatocysts or cysts in males and within germline cradles in females, from germline stem cell precursors called spermatogonia or oogonia, respectively (Nishimura et al., 2016). Histological examination of adult gonads showed that medaka ndrg1b mutants displayed typical female and male gonadal structure with all the different stages of germ cells; however, they had a lower production of gametes. In this regard, although a previous study has shown that type I proliferation is important to germline stem cell number in adult medaka gonads (Morinaga et al., 2007), here we showed that type II proliferation (cystic) is also crucial to gamete production, and consequently, to related reproductive parameters such as the number of spawned eggs, spermatozoa, and fertilization rate. Moreover, expression of ndrg1b in testis is restricted to the most peripheral region of the tubule, where A-type undifferentiated spermatogonia (SGa) are present (Iwai et al., 2006). This location of ndrg1b expression is in agreement with the reduction of cystic proliferation since it is not observed in B-type spermatogonia (SGb), which have already entered that proliferation.
Additionally, we observed that the reduction of cystic proliferation in ndrg1b mutants did not induce any sex reversal or modification in the sex ratio. These observations are in agreement with Nishimura et al., 2018 in figla, meioC, and dazl mutants in medaka, in which follicle formation is disrupted, germ cells are unable to commit to gametogenesis, and germ cells do not develop into EGSCs, respectively. All these mutants exhibited female secondary characteristics and ovary structures, demonstrating that the mechanism underlying sex reversal is manifested before germline cells exit from the status of stem cells/gonocytes and enter the gametogenic program (Nishimura et al., 2018). In contrast, medaka mutants that had an effect on type I proliferation, like the germ cell-deficient cxcr4 (Kurokawa et al., 2007) and also loss of amhrII (TGF-β pathway member) that display hyperproliferative germ cells (Morinaga et al., 2007; Nakamura et al., 2012), showed sex reversal. Interestingly, our results showed that the TGF-β pathway is not involved in the cystic division, and the increased proliferation in males is due to inhibition of TGF-β members involved in controlling germ cell type I proliferation, such as amh. Further studies are therefore necessary to elucidate the regulatory pathways of this type of proliferation.
An interesting observation in the present work was the lower spawn by normal females paired with ndrg1b mutant males. It has been established that sexual behavior is linked to reproductive success (Oshima et al., 2003), and, fortunately, the sexual behavior of medaka consists of a sequence of several actions that are easily quantified (Ono and Uematsu, 1957; Walter and Hamilton, 1970). We specifically observed differences in male mating behavior related to stimulation of female spawning and fertilization, such as the duration of quivering and number of convulsions; actions that seem to be conserved in vertebrates because ejaculation in male rats is accompanied by strong vibration and significant abdominal contraction (Qin et al., 2017). Sexual behavior has been found to be strongly controlled by the brain (Hiraki-Kajiyama et al., 2019; Mitchell et al., 2020; Yang and Shah, 2014) and levels of sex steroid hormones (Hiraki-Kajiyama et al., 2019; Munakata and Kobayashi, 2010). Here, it is important to note that in medaka the secondary sex characters are induced by sex steroid hormones (Kikuchi et al., 2019; Sato et al., 2008). ndrg1b mutants showed normal secondary sex characters in both males and females. Moreover, the level of the main androgen in fish, 11-KT, did not show differences between wt and ndrg1b mutants, thus establishing that sex steroid synthesis is not impaired in these animals. Regarding sexual behavior, in studies where the regulation of sex hormones was altered, changes in sexual behavior were primarily related to courtship, such as the frequencies of following, dancing, and latency from the first courtship (Hiraki-Kajiyama et al., 2019; Oshima et al., 2003). Additionally, when brain regulators were altered, such as arginine-vasotocin (Yokoi et al., 2015), oxytocin ligand (oxt) (Yokoi et al., 2020), and TN-GnRH3 (Okuyama et al., 2014), changes in courtship display and sexual motivation, latency to mating and number of courtships displays, and latency to mating were observed, respectively. None of these courtship actions changed in the male ndrg1b mutants, leading us to consider whether the specific mating actions of quivering and convulsions are exclusively related to the lower number of spermatogonia and spermatozoa in males. To our knowledge, only one other study has observed this loss of mating vigor in medaka. In it, successive multiple mating-induced depletion of sperm reserves decreased fertilization and mating rate, and even females responded by reducing clutch size (Weir and Grant, 2010). In this respect, this association warrants further investigation in vertebrates including brain, endocrine, and gonadal features.
Taken together, our data show that mutations of ndrg1b lead to disruption of germline stem cell cystic proliferation with a subsequent reduction in the number of gametes and an unexpected change in mating behavior of medaka. Altogether, these data are significant for three main reasons: (1) we showed that ndrg1b is involved in the regulation of cystic proliferation during different stages of medaka development (embryo, juvenile, and adult) in a TGF-β-independent manner; (2) we demonstrated that mutation of ndrg1b leads to changes in number of gametes, decreasing reproductive success for both sexes; and (3) ndrg1b mutation affected male mating behaviors in medaka without changing androgen production.
Materials and methods
Source of medaka
Request a detailed protocolAll experiments were performed with medaka (Oryzias latipes) (strain hi-medaka, ID: MT835) supplied by the National BioResource Project (NBRP) Medaka (http://www.shigen.nig.ac.jp/medaka/). Fish were maintained and fed following standard protocols for medaka (Kinoshita et al., 2012). Fish were handled in accordance with the Universities Federation for Animal Welfare Handbook on the Care and Management of Laboratory Animals (http://www.ufaw.org.uk) and internal institutional regulations. Fertilized medaka eggs were incubated in 60 mm Petri dishes with embryo medium (17 mM NaCl, 0.4 mM KCl, 0.27 mM CaCl2·2H2O, and 0.66 mM MgSO4; pH 7) until hatching and subsequently in 2 l water tanks to adults under a constant photoperiod (14L:10D) in a closed circulation water system at a controlled temperature (26 ± 0.5°C).
Sample collection
Request a detailed protocolSamplings were performed at different stages of medaka development: embryonic stages 35 and 39 (5 and 9 days post fertilization, respectively) and juvenile–adult stages 10, 20, and 80 dph, according to a previous description of medaka development (Iwamatsu, 2004). These stages are important for medaka gonadal development. Stage 35 corresponds to a sexually undifferentiated gonad in both sexes and only type I divisions (first mitotic proliferation). This stage precedes the beginning of type II division (cystic proliferation) in XX gonads (stages 36–39). Stage 39 corresponds to the maximum EGSC proliferation in XX embryos, and involves sexually dimorphic gonads, just at the time of hatching. At 10 dph, type II division is present only in XX gonads, while in XY gonads, it begins only at 20 dph. At 80 dph, animals have become sexually mature adults. For all sample collections, fish were euthanized by immersion in tricaine at 30 ± 50 mg/l and processed according to the technique to be used. To determine genotypic sex, DNA from the tail and head of each animal was analyzed by PCR to determine the presence of the dmy/dmrt1bY gene; ß-actin gene was used as a DNA loading control (Supplementary file 1A; Nanda et al., 2002). The PCR products were analyzed on 1% agarose gel.
Whole-mount RNA in situ hybridization
Request a detailed protocolWhole-mount RNA in situ hybridizations were performed as previously described (Nakamura et al., 2006). Fluorescein isothiocyanate (FITC)- and digoxigenin (DIG)-labeled probes were synthesized from the full-length medaka cDNA of ndrg1a, ndrg1b, and oct4 (stem cell marker also known as pou5f1; Wang et al., 2011) using pGEM-T Vector (Promega) linearized plasmid. Embryos at stage 35, and male brains and testis were fixed overnight in 4% RNAse-free paraformaldehyde at 4°C, permeabilized using 20 µg/µl proteinase K at room temperature (RT), and hybridized at 68°C overnight with either ndrg1a, ndrg1b, or oct4 DIG-labeled RNA probes, or at 66°C overnight with ndrg1b FITC-labeled RNA probe. Hybridized DIG probes were detected using an alkaline phosphatase–conjugated anti-digoxigenin antibody (1:2000; Roche) in the presence of nitro blue tetrazolium/5-bromo-4-chloro-3′-indolyphosphate substrates (Roche). Stained embryos were embedded in gelatin, cryostat sectioned at 14–16 µm thickness, and photographed. Hybridized FITC probes were examined under a fluorescence microscope.
RNA quantification by RT-qPCR
Request a detailed protocolEmbryos at stages 35 and 39, juveniles at 10 and 20 dph, and tissue of male brains and testis were used for gene expression analysis. For this purpose, the gonadal region of the body was immediately frozen in liquid nitrogen and subsequently stored at −80°C until RNA extraction. Once the sex of each individual was known by PCR, total RNA was extracted from a pool of five embryos, or individually from juveniles, using 300 µl of TRIzol Reagent (Life Technologies) following the manufacturer’s instructions. RNA from each sample (500 ng) was used to perform cDNA synthesis, following previous studies (Castañeda Cortés et al., 2019). Real-time PCR primers are listed in Supplementary file 1A. Gene-specific RT-PCR was performed using the SYBR green master mix (Applied Biosystem) and Agilent MX3005P Multiplex QPCR Real-time Thermal Cycler (Stratagene). The amplification protocol consisted of an initial cycle of 1 min at 95°C, followed by 10 s at 95°C and 30 s at 60°C for a total of 45 cycles. The subsequent quantification method was performed using the 2-ΔΔCt method (threshold cycle; assets; thermofisher.com/TFS-Assets/LSG/manuals/cms_040980.pdf) and normalized against reference gene values for ribosomal protein L7 (rpl7) (Zhang and Hu, 2007) or elongation factor 1 alpha (ef1α) (Hirayama et al., 2006).
TGF-β inhibitor treatment
Request a detailed protocolTo analyze whether TGF-β regulates ndrg1b expression, A83-01 (Sigma-Aldrich, USA), a selective TGF-β inhibitor was used (Tojo et al., 2005). A83-01 inhibits the phosphorylation of Smad2, thereby blocking the signaling of TGF-β type I receptor (Tojo et al., 2005). A83-01 was first dissolved in DMSO (3 mM) and subsequently diluted in embryo medium (17 mM NaCl, 0.4 mM KCl, 0.27 mM CaCl2·2H2O, and 0.66 mM MgSO4; pH 7) to obtain a solution of 3 μM. Fertilized medaka eggs were incubated in 70 mm Petri dishes with or without 3 µM A83-01 from stage 35 to stage 39 at 26°C (Figure 1—figure supplement 1). Freshly made dilutions were used, and the medium was changed every 24 hr. As control, a group of embryos were incubated only with 0.1% DMSO. The concentration used in this study was based on Tojo et al., 2005. After the incubation period, embryos were collected at stage 39 for sex genotyping, RT-qPCR, and cell quantification by histological analysis, as described in this article.
Generation of ndrg1b mutants using CRISPR/Cas9-induced mutagenesis
Request a detailed protocolCRISPR/Cas9 target sites were identified and designed using the medaka genome reference available on the Ensembl genome database (ENSORLG00000004785) and the CCTop-CRISPR/Cas9 target online predictor (CCTop, http://crispr.cos.uni-heidelberg.de/) (Stemmer et al., 2015). Two sequences 5'GG-(N18)-NGG3' in exon 6 of ndrg1b were identified. The selected target sites are shown in Fig. S1A and Fig. S2A. Each sgRNA was synthesized according to a protocol previously established by Ansai and Kinoshita, 2014. Briefly, the pair of inverse-complementary oligonucleotides was annealed and cloned into the pDR274 vector (Addgene #42250) in the BsaI cloning site. The modified pDR274 vectors were digested with DraI, and each ndrg1b-specific guide RNA (SgRNA_ndrg1b) was transcribed using the MEGAshortscript T7 Transcription Kit (Thermo Fisher Scientific). The synthesized sgRNAs were purified by RNeasy Mini kit purification (Qiagen). For the Cas9-RNA in vitro synthesis, the pCS2-nCas9n vector (Addgene #47929) was linearized with NotI, and capped Cas9 RNA was transcribed with the mMessage mMachine SP6 Kit (Thermo Fisher) following the manufacturer’s instructions and purified with the RNeasy Mini kit (Qiagen). Cas9-RNA (100 ng/µl) and SgRNA_ndrg1b (50 ng/µl) were co-injected into one-cell stage embryos as described previously (Kinoshita et al., 2000). Embryos injected only with cas9 mRNA were used as controls (wt). Microinjections were performed with a Nanoject II Auto-Nanoliter Injector (Drummond Scientific) coupled to a stereomicroscope (Olympus).
Biallelic CRISPR mutants screening and off-target analysis
Request a detailed protocolTo analyze the efficiency and specificity of the CRISPR/Cas9 system, genomic DNA was extracted with alkaline lysis buffer using 3 dpf embryos, as described previously (Ansai and Kinoshita, 2014), DNA was used as a template for PCR prior to HMA using the primers listed in Supplementary file 1A. Electrophoresis was performed on 10% acrylamide gels and stained with ethidium bromide (Ota et al., 2013). Multiple heteroduplex bands shown by HMA in PCR amplicons from each injected embryo were quantified as embryos with biallelic mutations, whereas single bands were quantified as non-edited embryos. The mutation rate was calculated as the ratio of the number of multiple heteroduplex bands shown in PCR amplicons from each Cas9-sgRNA-injected embryo to the sum of all embryos injected multiplied by 100 (n = 20/per sgRNA) (Ansai and Kinoshita, 2014). Sg1RNA_ndrg1b was selected for subsequent phenotypic analysis due to its greater efficiency. Additionally, potential off-target sites in the medaka genome for Sg1RNA_ndrg1b were searched for by using the CCTop-CRISPR/Cas9 target online predictor (Stemmer et al., 2015). All potential off-target sites identified were analyzed by HMA using the primers listed in Supplementary file 1A. The screening of indels was performed in F1 fish. Biallelic mutant adult (F0) medaka were mated with wt medaka. Genomic DNA was extracted from each F1 embryo for analysis of mutations by HMA as described previously (Supplementary file 1A). Mutant alleles in each embryo were determined by sequencing of the ndrg1b genomic DNA region. All phenotypic analyses were performed using positive ndrg1b biallelic mutants F0 injected with Cas9 and Sg1RNA_ndrg1b (SgN1b) and Cas9-injected individuals as controls (wt). Additionally, with the goal of analyzing KO individuals, an F2 generation was obtained by mating F1 fish harboring the same mutation (the Δ31, deletion of 31 bp) with each other; the obtained F2 individuals were analyzed by HMA genotyping using genomic DNA extracted from each fin clip of F2 80 dph fish as a template for PCR prior to first HMA using the primers listed in Supplementary file 1A, following the protocol previously established by Ansai and Kinoshita (Hatada, 2017). For reliable distinction between wt and KO fish (both homozygous show a single band pattern in first HMA), each PCR product from the first HMA was mixed with a wt template separately, then reannealed by heating at 95°C for 5 min and gently cooled to RT to perform the second HMA, where the wt will show only one band, while KO fish will show a multiple banding pattern similar to heterozygotes (Hatada, 2017; Figure 2—figure supplement 3).
Immunofluorescence analysis
Request a detailed protocolGonadal regions from individuals at stage 39 were processed, fixed in Bouin’s solution, embedded in paraffin, and sagittal sectioned at 5 µm. Sections were washed with 0.1 M phosphate-buffered saline (PBS pH 7.4) and blocked in 0.1 M PBS containing 0.5% bovine serum albumin (Sigma-Aldrich) and 0.5% Triton X-100 for 60 min before overnight incubation with primary antibody anti-OLVAS (1:200, rabbit, Abcam 209710) and anti-PCNA (1:200, mouse, Sigma P8825) at 4°C. After incubation, the sections were washed twice in PBS and incubated at RT for 90 min with Alexa Fluor 488-conjugated goat anti-rabbit IgG (Thermo Fisher Scientific, A-11008) and 594-conjugated goat anti-mouse IgG (Thermo Fisher Scientific, A32742) secondary antibodies at a dilution of 1:2000 in PBS. After incubation, sections were rinsed twice with PBS and mounted with Fluoromount mounting medium (Sigma-Aldrich) containing 4',6-diamidino-2-phenylindole (DAPI, 5 µg/ml, Life Technologies).
TUNEL assay
Request a detailed protocolThe presence of apoptosis at stage 39 was detected through the In situ Cell Death Detection Kit, Fluorescein (Roche). Samples fixed in Bouin’s solution, embedded in paraffin, and sagittal sectioned at 5 µm were treated according to the manufacturer’s manual, with a step of permeabilization with 0.1% Triton X-100, 0.1% sodium citrate in PBS 1× solution. After labeling the reaction for 1 hr at 37°C, sections were immunostained with OLVAS, as described above.
Histological analysis
Request a detailed protocolGonadal samples from juveniles at 20 (from whole body) and 80 dph (from gonadal tissue) were fixed in Bouin’s solution, embedded in paraffin, sectioned at 5 µm, and subjected to standard hematoxylin and eosin staining. For the TGF-β inhibitor treatment, stage 39 individuals were fixed in 4% buffered glutaraldehyde, embedded in Technovit (7100-Heraeus Kulzer), sectioned at 5 μm thickness, and stained with 0.1% toluidine blue to quantify the germ cells using a light microscope (Leica DM6000 BD, Leica Microsystems).
Cell quantification
Request a detailed protocolAll section photographs were taken using a ZEISS Axio Observer 7 Led colibri 7, Nikon Eclipse E600 and Nikon Digital Sight DSQi1Mc. Images were analyzed using FIJI software (https://imagej.nih.gov/ij/). Individual cells were counted manually with the Cell Counter plugin for FIJI. Type I and II germ cells were counted as previously described (Saito et al., 2007), where single isolated germ cells were counted as type I, while clusters with more than two germ cells were counted as germ cells undergoing type II division. Spermatogonia were counted based on the description by Iwasaki et al., 2009 and oocytes based on Seki et al., 2004 and Gay et al., 2018. All gonadal sections were counted for stage 39 individuals. For 20 and 80 dph individuals, three gonadal sections were counted, setting the distance between each section to avoid counting the same cell twice. Each section was counted two times in all experiments to reduce technical errors.
Sperm quantification
Request a detailed protocolSperm was collected from mutant and control adult males after immersion in tricaine at 30 ± 50 mg/l. Each male was placed on a foam rubber holder with the belly facing up; the urogenital pore was dried and sperm was collected with a micropipette while gentle pressure was applied from bilateral abdominal toward the anal opening using fine forceps. Prior to the analysis, the sperm was diluted 5000 times in cold Ringer’s solution (NaCl 0.75%, KCl 0.02%, CaCl2 0.02%, NaHCO3 0.002%) and kept at 4°C. Sperm counts were determined by hemocytometer (Bright-Line, Hausser Scientific) analysis under light microscope at ×200 magnification, performing three counting replicates per individual.
Evaluation of reproductive success
Request a detailed protocolTo evaluate reproductive success, we used three categories of sexually mature couples: control (wt female and male), mutant female (wt male and ndrg1b mutant female), and mutant male (wt female and ndrg1b mutant male). Each couple was acclimated for 5 days in a breeding tank to ensure and standardize their reproductive conditions. For 10 consecutive mornings, each couple’s fertilized and unfertilized eggs were carefully collected by hand, placed in a separate dish, and counted to calculate total spawned eggs, fertilization (number of fertilized eggs/number spawned eggs), and hatching (number of hatched larvae/number of fertilized eggs).
Sexual behavior assay
Request a detailed protocolSexual behavior assay was performed over 10 days using two categories of sexually mature couples, control and mutant male, mentioned above, which were allowed to acclimate for 5 days to ensure and standardize their reproductive conditions. On the remaining 5 days, the procedure was performed as previously described (Okuyama et al., 2014). Males and females were separated in the evening (6–7 PM) the day before the assay using a transparent plastic cup with small holes for water exchange. The following morning the mating pairs were placed together in a single transparent tank and their sexual behavior was recorded for 20 min using a digital video camera (Figure 5A). The recording order was randomly changed every day. Water inflow to the tanks was stopped during recording. Females that did not spawn or spawned without any mating behavior were excluded from the analysis. The following behavioral parameters were calculated from the video recordings: latency until first courtship display, latency from the first courtship display to the wrapping that resulted in spawning; number of wrappings before and after spawning, the duration of wrapping, and within wrapping time; and the duration of quivering and number of convulsions. Each action during sexual behavior was identified following the descriptions by Ono and Uematsu, 1957 and Walter and Hamilton, 1970 and analyzed twice by two different authors of the present study who did not know the category of the couple.
Blood levels of 11-KT in sexually mature males
Request a detailed protocolPlasma 11-KT concentrations were measured using an enzyme-linked immunosorbent assay kit (Cayman 582751) according to the manufacturer's instructions. Briefly, control and sgN1b sexually mature males (n = 7) were coupling with wt females (Figure 5—figure supplement 1A). Males and females were separated in the evening (6–7 PM) the day before the assay using a transparent plastic cup with small holes for water exchange, and then blood samples of anesthetized male individuals were collected directly from the caudal vein using a glass capillary tube (Royan et al., 2020). All fish were recovered after blood extraction. Steroid extractions were performed twice with ethyl acetate/hexane from 1 μl of whole blood and then reconstituted in 200 μl of assay buffer. All samples were assayed in duplicate, and hormone levels were determined based on a standard curve.
Statistical analysis
Request a detailed protocolValues are presented as mean ± standard error of the mean for continuous variables and as percentages for categorical variables. Fold change and statistical analysis of RT-qPCR quantifications were performed using FgStatistics software (http://sites.google.com/site/fgStatistics/), based on the comparative gene expressions method (Pfaffl, 2001). Statistical analyses were performed by using GraphPad Prism (GraphPad Software, San Diego, CA). Continuous variables were compared between two groups by the unpaired two-tailed Student’s t-test. If the F-test indicated that the variance differed significantly between groups, Welch’s correction to the Student’s t-test was employed. For more than two groups, continuous variables were compared by one-way analysis of variance, followed by Tukey’s multiple comparisons test, comparing the mean of each column with the mean of every other column. All differences were considered statistically significant when p<0.05.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files.
References
-
C. elegans anillin proteins regulate intercellular bridge stability and germline syncytial organizationJournal of Cell Biology 206:129–143.https://doi.org/10.1083/jcb.201310117
-
Sperm RNA elements as markers of healthSystems Biology in Reproductive Medicine 64:25–38.https://doi.org/10.1080/19396368.2017.1393583
-
Gonad morphogenesis in vertebrates: divergent means to a convergent endAnnual Review of Cell and Developmental Biology 25:457–482.https://doi.org/10.1146/annurev.cellbio.042308.13350
-
Molecular functions of the iron-regulated metastasis suppressor, NDRG1, and its potential as a molecular target for Cancer therapyBiochimica Et Biophysica Acta (BBA) - Reviews on Cancer 1845:1–19.https://doi.org/10.1016/j.bbcan.2013.11.002
-
The regulatory network controlling the proliferation-meiotic entry decision in the Caenorhabditis elegans germ lineCurrent Topics in Developmental Biology 76:185–215.https://doi.org/10.1016/S0070-2153(06)76006-9
-
BookMethods in Molecular BiologyIn: Hatada I, editors. Genome Editing in Animals: Methods and Protocols. Springer. pp. 175–188.https://doi.org/10.1007/978-1-4939-7128-2
-
Temperature-dependent growth rates and gene expression patterns of various medaka Oryzias latipes cell lines derived from different populationsJournal of Comparative Physiology B 176:311–320.https://doi.org/10.1007/s00360-005-0053-8
-
Mutation of gonadal soma-derived factor induces medaka XY gonads to undergo ovarian developmentBiochemical and Biophysical Research Communications 467:109–114.https://doi.org/10.1016/j.bbrc.2015.09.112
-
Structural components of the synaptonemal complex, SYCP1 and SYCP3, in the medaka fish Oryzias latipesExperimental Cell Research 312:2528–2537.https://doi.org/10.1016/j.yexcr.2006.04.015
-
Stages of normal development in the medaka Oryzias latipesMechanisms of Development 121:605–618.https://doi.org/10.1016/j.mod.2004.03.012
-
Two states of active spermatogenesis switch between reproductive and non-reproductive seasons in the testes of the medaka, Oryzias latipesDevelopment, Growth & Differentiation 51:521–532.https://doi.org/10.1111/j.1440-169X.2009.01114.x
-
Activity of the medaka translation elongation factor 1alpha-A promoter examined using the GFP gene as a reporterDevelopment, Growth and Differentiation 42:469–478.https://doi.org/10.1046/j.1440-169x.2000.00530.x
-
BookMedaka: Biology, Management, and Experimental ProtocolsAmesUnited States: John Wiley & Sons, Ltd.https://doi.org/10.1002/9780813818849
-
The Iron-Regulated metastasis suppressor NDRG1 targets NEDD4L, PTEN, and SMAD4 and inhibits the PI3K and ras signaling pathwaysAntioxidants & Redox Signaling 18:874–887.https://doi.org/10.1089/ars.2011.4273
-
Expression of NDRG1, a differentiation-related gene, in human tissuesHistochemistry and Cell Biology 118:399–408.https://doi.org/10.1007/s00418-002-0460-9
-
BookComparative Vertebrate ReproductionSpringer Science & Business Media.https://doi.org/10.1007/978-1-4615-4937-6
-
Comparative aspects of animal oogenesisDevelopmental Biology 231:291–320.https://doi.org/10.1006/dbio.2000.0120
-
Phosphorylation of NDRG1 is temporally and spatially controlled during the cell cycleBiochemical and Biophysical Research Communications 411:227–234.https://doi.org/10.1016/j.bbrc.2011.06.092
-
Endocrine control of sexual behavior in teleost fishGeneral and Comparative Endocrinology 165:456–468.https://doi.org/10.1016/j.ygcen.2009.04.011
-
Fused regulates germline cyst mitosis and differentiation during Drosophila oogenesisMechanisms of Development 123:197–209.https://doi.org/10.1016/j.mod.2006.01.001
-
A structurally and functionally common unit in testes and ovaries of medaka (Oryzias latipes), a teleost fishSexual Development : Genetics, Molecular Biology, Evolution, Endocrinology, Embryology, and Pathology of Sex Determination and Differentiation 10:159–165.https://doi.org/10.1159/000447313
-
A new mathematical model for relative quantification in real-time RT-PCRNucleic Acids Research 29:45e–45.https://doi.org/10.1093/nar/29.9.e45
-
Mechanisms guiding primordial germ cell migration: strategies from different organismsNature Reviews Molecular Cell Biology 11:37–49.https://doi.org/10.1038/nrm2815
-
Gonadectomy and blood sampling procedures in the small size teleost model japanese medaka (Oryzias latipes)Journal of Visualized Experiments 1:62006.https://doi.org/10.3791/62006
-
Sex differentiation of germ cells in the teleost, Oryzias latipes, during normal embryonic developmentJournal of Embryology and Experimental Morphology 28:385–395.https://doi.org/10.2108/zsj.20.1231
-
Fish full life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes)Environmental Toxicology and Chemistry 23:774–781.https://doi.org/10.1897/03-26
-
Genetic control of gonadal sex determination and developmentTrends in Genetics 35:346–358.https://doi.org/10.1016/j.tig.2019.02.004
-
Identification of pluripotency genes in the fish medakaInternational Journal of Biological Sciences 7:440–451.https://doi.org/10.7150/ijbs.7.440
-
Novel thiosemicarbazones inhibit Lysine-Rich carcinoembryonic Antigen-Related cell adhesion molecule 1 (CEACAM1) Coisolated (LYRIC) and the LYRIC-Induced Epithelial-Mesenchymal transition via upregulation of N-Myc Downstream-Regulated gene 1 (NDRG1)Molecular Pharmacology 91:499–517.https://doi.org/10.1124/mol.116.107870
-
TGF-β family signaling in the control of cell proliferation and survivalCold Spring Harbor Perspectives in Biology 9:a022145.https://doi.org/10.1101/cshperspect.a022145
Article and author information
Author details
Luisa F Arias Padilla

Funding
Agencia Nacional de Promoción Científica y Tecnológica (0366/12)
- Juan I Fernandino
Agencia Nacional de Promoción Científica y Tecnológica (2501/15)
- Luisa F Arias Padilla
Fundação de Amparo à Pesquisa do Estado de São Paulo (14/07620-7)
- Ivana F Rosa
- Rafael H Nóbrega
Fundação de Amparo à Pesquisa do Estado de São Paulo (18/10265-5)
- Ivana F Rosa
- Rafael H Nóbrega
Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET and São Paulo Research Foundation International Cooperation Grant D 2979/16)
- Ricardo S Hattori
- Juan I Fernandino
São Paulo Research Foundation (CONICET and São Paulo Research Foundation International Cooperation Grant D 2979/16)
- Ricardo S Hattori
- Juan I Fernandino
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Tech. Gabriela C López and Dr. Leandro A Miranda (INTECH) for helping with histological preparations. We also thank Dr. Tania Rodriguez for technical support and Tech. Javier Herdman (INTECH) for fish handling. We are grateful to NBRP Medaka (https://shigen.nig.ac.jp/medaka/) for providing HNI (Strain ID: MT835). We thank Dr. John M Murray (Biodesign Center for Mechanisms of Evolution, Arizona State University) for critical reading of the manuscript. This work was supported by the Agencia Nacional de Promoción Científica y Tecnológica Grant 0366/12 and 2501/15 (to JIF). RHN and IFR were supported by São Paulo Research Foundation (FAPESP), Brazil (Grants 14/07620-7 and 18/10265-5), CONICET and Sao Paulo Research Foundation (FAPESP), and International Cooperation Grant D 2979/16 (to JIF and RSH). LFAP, DCCC, and ODMA were supported by a PhD scholarship from the National Research Council (CONICET). JIF is a member of the Research Scientist Career at the CONICET.
Copyright
© 2021, Arias Padilla et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,651
- views
-
- 208
- downloads
-
- 13
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 13
- citations for umbrella DOI https://doi.org/10.7554/eLife.62757
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cystic proliferation of germline stem cells is necessary to reproductive success and normal mating behavior in medaka
eLife 10:e62757.
https://doi.org/10.7554/eLife.62757




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}